Časť 2. Pôvod duše a kultúry
Časť 2. Pôvod duše a kultúry
Darwinistická myseľ
Darwinistická myseľ
Jednou z kľúčových koncepcií mémológie je založená na darwinisticko-ekologickej predstave mysle, alebo ak chceme duše. Podľa nej je myseľ ekosystém mémov, ktoé majú medzi sebou zložitý systém symbiotických a konkurenčných vzťahov.
Z darwinistického náhľadu taktiež vyplýva, že všetky mémy, tak ako znaky organizmov, či iné darwinistické entity, majú svoju evolučnú minulosť, nezjavujú sa v mysli zčista-jasna zázrakom. Podstatnú časť našej mysle tvoria kultúrne mémy, a tie sú prevažne lokalizované v mozgu. Zdôrazňujem PREVAŽNE, pretože miera podieľania sa iných častí tela na tvorbe „pocitu“, základu to každého mémplexu, je dosť nepreskúmaná a môže byť často podstatná.
Mozog je darwinistické prostredie, pretože v ňom prebieha kompetícia mémov, tvoria sa mutácie, teda variácie mémov, prebieha prírodný výber, ktorý má dve formy:
- podobu zabúdania a
- neschopnosti mému prejsť z tzv. krátkodobej do dlhodobej pamäte.
Z charakteru neuronálnej siete mozgu vyplýva, že mém je štruktúralne uložený v synapsách medzi neurónmi a zdá sa ešte, že aj v určitých vnútrobunečných mechanizmoch neurónov, napríklad v miere intenzity produkcie mRNA molekúl, ktoré sa podieľajú na tvorbe synaptických prenosov. Z takéhoto hľadiska je pre mém dôležité, v koľkých synaptických, či vnútrobunečných reláciách sa vyskytuje. Darwinisticky povedané: koľko má potomkov.
Prakticky žiadna informácia v mozgu nie je uložená osamote, ako v pamäti počítača. Neurón je miesto, kde sa stretáva mnoho informácii z mnohých miest mozgu. Mém preto neexistuje osamotene, je z asociatívnej podstaty mozgu vždy v symbióze s inými mémami mozgu. Pravda, pokiaľ sa mu podarí túto symbiózu vytvoriť. To má niekoľko koncepčných dôsledkov.
Prvý dôsledok je chronologicko-topologický. Štruktúra ekosystému mémov v mozgu vzniká ako hierarchický systém v procese ontogenézy mozgu postupne. Mozog sa rodí s už daným vrodeným ekosystémom mémov, ku ktorým mémy prichádzajúce prostredníctvom zmyslov a iných vstupov musia vytvárať mémplexy. Týmto sa mení ekosystémové usporiadanie mozgu a vznikajú nové pravidlá pre následne prichádzajúce mémy. Tento postup sa neustále cyklicky opakuje počas celej ontogenézy mozgu. Ide o typický darwinistický proces, pri ktorom sa nevyvíjajú len darwinistické entity, ale súčasne aj pravidlá prírodného výberu nad nimi v ďalšom kole.
Z pohľadu mémov je dôležité, aký dlhý čas prebieha ontogenéza mozgu. Ontogenéza mozgu šimpanza trvá zhruba dva roky. Za ten čas by mal byť schopný šimpanz mentálne ovládnuť svoje prostredie. Následne sa mozog morfologicky a vývojovo ukončí, stáva sa dospelým ako ostatné časti organizmu. U sapienta však tento proces nenastane [1]. Náš mozog teda ostáva morfologicky a vývojovo meukončený, mláďací do smrti, čo je zjavne jednou z hlavných príčin jeho fenotypovej plasticity. Môže sa prakticky učiť, replikovať kultúrne mémy, skoro do smrti.
Ako možno potom vysvetliť zjavnú skutočnosť, že čím je jedinec starší, tým je menej schopný meniť názory, učiť sa nové sociálne schopnosti a reagovať na meniace sa podmienky v spoločnosti? Napríklad staršie generácie majú neprekonateľné problémy ovládať produkty technologického vývoja, začlenňovať sa do meniacej spoločenskej komunikácie, ovládať nové pracovné návyky, atď. Časť odpovede sa iste nachádza v zhoršujúcej sa fyziológii neurónov. To je iste pravda, čo sa týka veku nad 60 rokov. Pravdou však je, že spomínaná znížená schopnosť replikácie nových generácií kultúrnych mémov sa začína prejavovať už oveľa skôr. Jedinec po tridsiatke začína byť názorove konzervatívny, menej ochotný k zásadným zmenám v modeli prostredia, ktorý je reprezentovaný ekosystémom mémov v jeho mozgu.
Myslím, že odpoveď na túto skutočnosť treba hľadať v hierarchickej topológii ekosystému mémov, ktorú si môžeme názorne reprezentovať topológiou stromu.. Mémy, ktoré prídu ako prvé, vytvárajú hlavné vetvy, na tých sa budujú závislé vetvy a na tých ešte závislejšie atď. V podstate je to hierarchia mémov do rôznych mémplexov a ekosystémové vzťahy medzi rôznymi mémplexami. Takto vybudované štruktúra postupne stále menej umožňuje vytváranie zásadných zmien. Nové kultúrne mémplexy nenachádzajú vhodných symbiontov.
Druhý dôsledok je ekologický, teda nemožnosť vytvárania izolovaných ostrovov mémov v mozgu. Každý nový podnet z prostredia, každý mémplex, ktorý sa pokúšať do mozgu vreplikovať, musí mať už svoju podstatnú v mozgu vytvorenú a iba pre niektoré mémy musí mozog vytvárať nové štruktúry. To je obrovská výhoda mozgu, pretože napríklad nemusí na každý mémplex vytvárať nové štruktúry, čím sa enormne šetria zdroje. Napríklad každá tvár je identfikovateľná určitými čiarami, preto tvar týchto čiar je jeden mém (mémplex?), ku ktorému sa postupne pridávajú iba detaily pre konkrétne osoby [2].
Tretí dôsledok tkvie v ekosystémovom chápaní mozgu. Mozog nemôžeme deliť na časti, z ktorých každá koná autonómne. Podobne ako v biosystémoch nie je možné popísať správanie organizmu ako vyčlenené z prostredia, a teda z ekosystémových vzťahov, tak aj žiaden mém, či mémplex v mozgu nie popísateľný bez jeho ekosystémových vzťahov. Súčasťou každej analýzy správania jedinca je preto nutnosť odhaľovať tieto ekosystémové zákonitosti. Je bezpočet štúdií, ktoré toto dokazujú. Napríklad pohlavne deteminované riešenie morálnych dilem, ktoré je závisle na vrodených mémplexoch pre empatiu a vyhodnocovanie rizík [3]. Je tak isto systémovo nesprávne deliť myseľ na funkčné celky, napríklad na vedomie a nevedomie.
Zdroje a literatúra:
- Bufill, E., Agustí, J., & Blesa, R. (2011). Human neoteny revisited: The case of synaptic plasticity. American Journal of Human Biology : The Official Journal of the Human Biology Council. doi: 10.1002/ajhb.21225
- Quiroga, R. Q., Fried, I., & Koch, C. (2013). Brain Cells for Grandmother. Scientific American, 308, 30 – 35. doi:10.1038/scientificamerican0213-30
- Friesdorf, R., Conway, P., & Gawronski, B. (2015). Gender Differences in Responses to Moral Dilemmas: A Process Dissociation Analysis. Personality and Social Psychology Bulletin, 41(5), 696–713. doi:10.1177/0146167215575731
Význam mozgu
Význam mozgu
Evolučný pohľad na svet je v princípe historizujúci, preto na akúkoľvek stávajúcu skutočnosť nazerá ako na bod v reťazci príčin a následkov. V darwinistickom zmysle duša nemôže vzniknúť náhle, bez nejakej predchádzajúcej formy, ale ako transformácia niečoho predchádzajúceho, transformácia, ktorá umožňuje organizmu prispôsobiť sa zmenám prostredia. Zrejme nebude pre nikoho prekvapením, že to, čo pomenúvame dušou, budeme väčšinou lokalizovať do orgánu, ktorému hovoríme mozog.
Ako každý orgán v tele, aj mozog plní určitú funkciu, na ktorú ho evolučný vývoj formoval desiatky miliónov generácií predkov. Aby sme túto funkciu správne popísali, musíme si uvedomiť, že každý organizmus aby dokázal prežiť vo svojom prostredí, musí byť naň funkčne prispôsobený, čo inými slovami znamená, že musí v sebe obsahovať nejaký funkčný model toho prostredia.
Čo si môžeme predstaviť pod funkčným modelom prostredia? Sú to nástroje, ktoré organizmus má, na riešenie situácií, do ktorých sa v prostredí dostáva. Ak musí napríklad vyhľadávať potravu, musí mať na to zmysly, pohybový aparát, rozoznávacie nástroje a všeličo iné. Tieto nástroje sú funkciami, ktoré organizmus obsahuje, funkciami, ktoré plnia úlohy dané prostredím. Sú to teda funkcie odrážajúce vzťah organizmu k prostrediu, a tvoria jeho funkčný model.
Dlhé obdobie vývoja života na Zemi bol tento model realizovaný pevným zápisom v genóme organizmov. Ak genóm jednotlivca sformoval správanie organizmu v prostredí menej presne, tak tento jedinec mal menej potomkov, alebo dokonca zahynul bez akýchkoľvek potomkov. Ak mal vrodený výhodnejší variant modelu prostredia, tak sa tento model v populácii rozšíril. Tomuto modelovaniu prostredia môžeme hovoriť genetické.
Pre náš výklad je dôležité si uvedomiť, že genetické modelovanie prostredia má tú nevýhodu, že prispôsobovanie sa organizmov zmenám v prostredí je generačné, čiže ak v prostredí nastanú zmeny, až nasledujúce generácie sú schopné ich vo svojej genetickej variabilite odraziť. Platí jednoduchá logika: ak je jedinec vopred (vrodene) prispôsobený zmene v prostredí, tak prežije.
Preto je hlavnou stratégiou takýchto druhov organizmov (napríklad baktérií) veľkosť populácie. Čím rýchlejšie a vo väčšom počte sa druh organizmov rozmnožuje, tým má väčšiu vnútornú variabilitu modelu prostredia, a teda má väčšiu šancu na druhové prežitie. Rýchlosť replikácie je však spojená s veľkými energetickými výdajmi, preto je genetické modelovanie prostredia výhodné pre jednoduché organizmy.
Napríklad ak podrobíme populáciu baktérií pôsobeniu antibiotika, ktoré rozkladá bunkovú membránu baktérie, tak väčšina populácie, ktorá má štandardne tvorenú bunkovú membránu, zahynie. Antibiotikum je zmena v prostredí, na ktorú baktéria nebola vrodene prispôsobená. Inými slovami: nemá vrodený model prostredia, v ktorom existujú antibiotiká. Lenže platí variabilta v populácii a niektoré baktérie majú trocha inak tvorenú membránu. Táto (pôvodne možno nevýhodná) variácia je iný vrodený model prostredia, ktorý zrazu nadobudne význam, a baktérie s týmto modelom založia novú populáciu.
Ak sa má organizmus vo svojom prostredí cielene pohybovať za zdrojmi (napríklad za potravou), tak potrebuje oveľa zložitejší vnútorný model prostredia, ako organizmus, ktorý na zdroje čaká. Zrejme preto sa postupne vyvinula najprv nervová a potom centrálna nervová sústava, ktorá umožňuje reagovať na meniace sa prostredie počas života jedinca lepšie, ako čisto statický vrodený systém. Dôsledkom vývoja centrálneho nervového systému je aj jeho súčasť — mozog. Mozog umožňuje vytvárať model prostredia na základe skúsenosti žijúceho jedinca. Jedinec tak pozná mapu svojho prostredia, rozloženie zdrojov potravy, partnerov na párenie, predátorov v okolí a mnohé iné prvky prostredia, ktoré sú individuálne, a preto nie je možné organizmus na ne vopred pripraviť genetickým modelovaním. Toto modelovanie budeme nazývať skúsenostné.
Je dôležité si uvedomiť, že z darwinistického hľadiska nejde o nič prevratne nové, pretože ide stále o ten istý princíp: malými adaptívnymi zmenami z generácie na generáciu sa postupne vytvoril prechod od druhového modelovania prostredia ku skúsenostnému.
Hlavná výhoda modelu, ktorý sa získava skúsenosťou je možnosť tzv. prediktívneho rozhodovania. To neznačí nič iné, ako to, že orgán skúsenostného modelovania prostredia — mozog — môže pri svojom rozhodovaní testovať možné rozhodnotia najprv na modeli prostredia, a až potom sa rozhodnúť. Lakonicky to vyjadril filozof Karl Popper (parafrázujem): Je oveľa výhodnejšie ak zahynie hypotéza o svete ako jej nositeľ. Napríklad je rozdiel ak rovno prejdem v teréne cez nejaké nebezpečné miesto, kde na mňa čaká predátor, alebo si pred pohybom predstavím možnosť priameho prechodu (hypotéza, že terén je voľný), alebo cez to miesto v teréne radšej nepôjdem (hypotéza, že tam predátor môže byť) a radšej pôjdem bezpečnejšou, hoci dlhšou trasou.
Mozog teda nielenže môže ukladať skúsenosť jedinca o prostredí do modelu tohoto prostredia, ale tiež vytvárať alternatívy tohoto modelu v prediktívnych variantách.
Mozog ako umwelt
Mozog ako umwelt
Každý organizmus nie je entitou samou osebe, ale je súčasťou celku — prinajmenšom ekosystému. Jeho súvislosť s celkom musí byť nejako
odrazená v štruktúre jeho vnútra. Musí sa teda skladať z častí, ktoré spoluvytvárajú jeho integritu a organizáciu a z častí,
ktoré určujú jeho vzťah k prostrediu, jeho znalosť prostredia. Túto predstavu formuloval na začiatku 20.stor Jakob von Uexküll [1] v koncepcii organizmu ako jednoty innenweltu (vnútra, vnútorného sveta) a umweltu (začlenenia, vnorenia do okolia) každého organizmu.
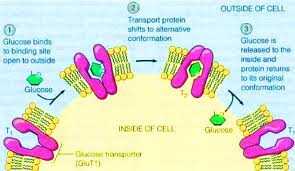 Umweltom bunky sú všetky jej znalosti o svete. Ak sa živí glukózou, tak do jej umweltu patrí zodpovedajúci receptor na bunkovej membráne, ktorý rozozná glukózu, naviaže sa na ňu a dopraví ju do vnútorného prostredia bunky. Ak na receptor dopadne iná molekula, povedzme kyseliny métanovej, tak ju bunka nepozná, nepatrí do jej umweltu. Do jej umweltu pochopiteľne nepatrí ani to, že jej reakcie skúmame v Petriho miske pod mikroskopom.
Umweltom bunky sú všetky jej znalosti o svete. Ak sa živí glukózou, tak do jej umweltu patrí zodpovedajúci receptor na bunkovej membráne, ktorý rozozná glukózu, naviaže sa na ňu a dopraví ju do vnútorného prostredia bunky. Ak na receptor dopadne iná molekula, povedzme kyseliny métanovej, tak ju bunka nepozná, nepatrí do jej umweltu. Do jej umweltu pochopiteľne nepatrí ani to, že jej reakcie skúmame v Petriho miske pod mikroskopom.
Ak sa na vec pozrieme morfologicky, tak vidíme, že receptor glukózy má časť, ktorá viac-menej presne zapadá na molekulu glukózy (viď fialovo označené časti na obrázku). Tvar molekuly glukózy je obsiahnutý v tvare jej receptora, je teda skopírovaný do umweltu bunky. Môžeme naň nazerať ako na replikátor, ktorý sa úspešne replikuje z prostredia do bunky tlakom prírodného výberu. Iný tvar molekuly má menšiu šancu sa do bunky zreplikovať, bunka, ktorá replikuje iné tvary tam, kde má byť tvar glukózy zahynie.
Na tomto jednoduchom príklade vidíme, že umwelt organizmu je tvorený úspešnými replikátormi prostredia. Pokojne ich nazvime mémy. Mémy samozrejme vyťažujú zdroje organizmu. Svet je plný tvarov, mémov, nie každý má rovnakú šancu uspieť v replikácii do umweltu organizmu. Bunka o Petriho miske nevie, pretože to nijako k svojmu životu nepotrebuje, prírodný výber ju nepripravil na takú situáciu, a preto molekulárna štruktúra sklenenej nádoby nemá šancu sa do umweltu bunky zreplikovať.
Mnohobunkové organizmy si vyvinuli schopnosť špecializácie buniek na rôzne zložky umweltu, bunky sa podľa špecializácie združujú do orgánov. Každý orgán sa nešpecializuje iba na určité zložky innerweltu, napríklad pečeň na metabolickú premenu látok, ale aj na časť umweltu, v prípade pečene na znalosť toho, aké molekuly prostredia sú vhodné pre metabolizmus jej a celého organizmu.
Pre organizmy, ktoré musia aktívne vyhľadávať potravu a partnerov na rozmnoženie sa umwelt musí obohatiť o priestorové, časové, sociálne a mnohé ďalšie mémy prostredia. Zrejme z tých istých dôvodov, z akých sa vytvorila orgánová koncentrácia bunkovej špecializácie pre iné zložky umweltu, sa pre spomínané aktívne znalosti prostredia vytvoril mozog.
Mozog je orgán na uloženie umweltu mnohobunkových organizmov.
Mozog, ako sa zväčša asi všetci zhodnú, je tvorený sieťou neurónov. Obsah informácie o prostredí je tvorený štruktúrou prepojení medzi neurónmi. Táto štruktúra je u jednoduchších organizmov, ako napríklad u hmyzu, definitívna, teda vytvorená počas ontogénezy jedinca vzájomným pôsobením dedičnej štruktúry DNA a epigenetických vplvov od vajíčka po dopselého jedinca.
Výhodou definitívnej štruktúry mozgu je jeho nízka spotreba zdrojov a pomerne jednoduchá ontogenéza. Nevýhodou je malá pružnosť. Prostredie sa môže meniť a nie každá konfigurácia mémov definitívneho mozgu je rovnako vhodná na prežitie v konkrétnom prostredí. Jedinci s vhodnejšou konfiguráciou mémov prežijú a majú viac potomkov ako tí, ktorých mémy až také vhodné nie sú.
Zrejme preto sa u stavovcov vyvinul pružný mozog ako nástavba nad starším mozgom pevným.
- Uexküll, Jakob. von. (1909). Umwelt und innenwelt der tiere (p. 276). Verlag von Julius Springer, Berlin. Retrieved from http://archive.org/details/umweltundinnenwe00uexk
Sebecké replikátory
Sebecké replikátory
Pripomeňme si základné aspekty Darwinovej evolučnej teórie:
- Potomkovia dedia podobu svojich predkov — druhová stabilita .
- Potomkovia síce dedia podobu svojich predkov, predsa len sa však v detailoch líšia — individuálna variabilita, diverzita .
- Vždy existuje viac potrieb využitia zdrojov, ako možností uspokojiť tieto potreby, alebo inak: vždy sa rodí viac potomkov ako môže prežiť — konkurencia .
- Odlišnosť potomkov spôsobuje rozdielnu schopnosť prežiť v súťaži o obmedzené zdroje — selekcia , alebo prírodný výber .
Z
pohľadu darwinizmu sú všetky znaky organizmu, či už ide o fyzické znaky
alebo prvky správania, adaptáciami na selekčný tlak prostredia. Od
počiatku bola táto teória problematická v otázke, ktoré časti prírody sú
vlastne vystavené tlaku prírodného výberu: druhy, či jednotlivci? Až
genetika priniesla do veci zreteľnejšie riešenie, ktoré vyústilo v tzv.
génocentrický model evolúcie, ktorý je v súčasnosti prevažne uznávaný.
Považujem za nevyhnutné na tomto mieste uviesť, že ja patrím k tej
menšine, ktorá nepovažuje génocentrický model evolúcie za dostatočný,
ale pre účely tohoto cyklu budem pre jednoduchosť vychádzať z neho.
Génocentrický
model evolúcie predpokladá, že jednotkou prírodného výberu je gén,
ktorý je dnes chápaný ako kúsok informácie zaznamenanej na molekule DNA,
alebo RNA. Gén je vlastne predpis, pomocou ktorého sa v bunke vyrába
konkrétna bielkovina (proteín). No a z proteínov je vlastne poskladaný
celý živý svet tak ako z kociek Lega.
V génocentrickom modeli nesúťažia o prežitie organizmy ale gény, čo je popísané nasledovnou schémou:
- Gén je informácia , zapísaná vo forme molekuly.
- Gén je molekula, ktorá má schopnosť sebaopakovania — replikácia. .
- Replikácia prebieha s rozličnou presnosťou — variabilita .
- Potomkovia pôvodného génu súťažia o zdroje. Zdrojmi sú predovšetkým organické molekuly, z ktorých sa poskladajú ďalší potomkovia génov — konkurencia .
- Tie gény, ktoré sú schopné získavať zdroje rýchlejšie, či sa naraz kopírovať vo viacerých kópiáchť, majú viac potomkov a postupne v populácii prevážia — selekcia, prírodný výber .
Tejto myšlienkovej schéme sa populárne hovorí
sebecký gén
. Teoreticky ju spracoval George C. Williams a spopularizoval ju Richard Dawkins, ktorý jej dal aj ten názov.
Z koncepcie sebeckého génu vyplýva, že organizmy sú nástroje (
vehicles
) génov, na presadenie ich sebeckosti. Gény si v priebehu vývoja vytvorili telá, mozgy, svaly na presadenie svojej sebeckosti.
Rovnako
však z tejto teórie vyplýva aj informačná podstata génu. Gén je
predovšetkým informácia, ktorá sebe vlastným spôsobom organizuje
molekuly do určitého usporiadania. Aby sme zvýraznili túto skotočnosť,
tak informačnú podstatu génu nazveme
replikátor
.
Replikátor je usporiadanie niečoho (v prípade génu usporiadanie
molekúl), ktoré má schopnosť opakovať sa, rozmnožiť — zreplikovať.
Aby
túto skutočnosť R. Dawkins dostatočne zdôraznil, hľadal v prírode iný
príklad na replikátor. Všimol si, že určité kultúrne javy, napríklad
móda, alebo vtipy sa opakujú, replikujú od sapienta k sapientovi. Sú to
svojou povahou informácie — replikátory. Nazval ich
mémy
, z gréckeho miméma (napodobniť), zrejme preto aby sa názov foneticky podobal na gén. Bližšie v knihe R. Dawkinsa
Sebecký gén.
Od toho momentu môžeme datovať zrod koncepcie, ktorá má potenciál darwinisticky vysvetliť dušu a kultúru sapientov bezo zvyšku.
Replikátory duše
Replikátory duše
V kapitole Význam mozgu som ukázal, že mozog je pre organizmus orgánom modelovania prostredia. Pre sociálne organizmy je prostredím nielen životné prostredie, ale aj sociálne prostredie. Z pohľadu mozgu je to jedno: prostredie ako prostredie. Aj sociálne prostredie má svoje zákonitosti, a tie treba vo forme skúsenosti uložiť do modelu, v rámci ktorého môže organizmus prijímať optimálne riešenia svojich životných situácií.
Čo je to vlastne skúsenosť? Majme ľubovoľný predmet, napríklad jablko. Jablko má tvar, zloženie, vlastnosti. Na jablko môžeme nazerať ako na štruktúru, usporiadanie, informáciu. Jablko je súčasť prostredia. Ak prídeme do styku s jablkom, reagujeme naň. Pokiaľ sme jablko dovtedy vôbec nepoznali, uložíme si do mozgu časť štruktúry „jablkovej“ informácie, teda časť usporiadania prostredia do mozgu. Pochopiteľne, do mozgu si neuložíme celé jablko, iba tú časť informácie, ktorá nám stačí na jeho identifikovanie. Napríklad obraz jablka, a jeho chuť. V našom funkčnom modeli prostredia v mozgu pribudli ďalšie asociácie, ďalšie prvky, ďalšia skúsenosť.
Ako som v kapitole Sebecké replikátory uviedol, súčasná génocentrická koncepcia evolúcie chápe gén ako element prírodného výberu. Gén nechápe primárne ako molekulu, ale ako informáciu, replikátor, ktorá usporiadava molekuly vo svoj sebecký prospech. Ak sa vrátime k nášmu príkladu s jablkom, tak vidíme, že z jablka ako neoddeliteľnej súčasti prostredia, ako usporiadania časti prostredia, teda informácie, sa ako skúsenosť zreplikovala časť toho usporiadania (obraz, chuť, vôňa, pocit spokojnosti, či odporu,…) do modelu prostredia v našom mozgu.
Informácia je nemateriálna, je to vždy vzťah medzi elementami, ktoré môžu byť materiálne. Na informácii nie je dôležitá jej podstata, ale jej štruktúra. Ak sa z prostredia do modelu prostredia zreplikuje nejaká informácia, tak v skutočnosti sa usporiadanie niečoho, v tomto príklade časti usporiadania prostredia — jablka — skopíruje do modelu prostredia, v tomto prípade do neuronálnej štruktúry mozgu. Informácia, ktorá formovala časť prostredia — jablko — teraz sformovala väzby v neuronálnej sieti.
To isté si skúsme vyjadriť pre názornosť na nasledovnom zobecňujúcom onrázku.Vidíme tam ako sa v prostredí reprezentovanom sieťou jeho prvkov (entít) časť prostredia (napríklad charakteristika jablka) skopíruje (zreplikuje) do modelu prostredia (napríklad mozgu), ktorý je reprezentovaný žlto podfarbeným výsekom prostredia.
 |
|
Replikácia skúsenosti
|
Pozrime
sa však na ten istý proces z darwinistického pohľadu. Ak sa nejaká
informácia skopírovala, zreplikovala, tak ide o evolučný proces a my ho
môžeme darwinisticky popísať. Táto informácia má predkov a potomkov. Ten
istý predmet, napríklad to jablko, sa môže dostať do kontaktu s
viacerými replikačnými nástrojmi — mozgami. Informácia bude teda mať
viac potomkov. Každý z týchto potomkov bude súčasne druhovo rovnaký, veď
vždy ide o to isté jablko, zároven však bude vždy trocha iný, pretože
každý z mozgov, ktoré ho ako skúsenosť zreplikovali má inú evolučnú
históriu, a teda tú istú informáciu z prostredia zreplikuje v trocha
inej štruktúre.
Zadefinujme si mém ako taký element usporiadania prostredia (informácie), ktorý sa z prostredia do modelu prostredia (napríklad mozgu) zreplikuje ako nedeliteľný celok.
Takýmto mémom bude v našom príklade napríklad tvar jablka, chuť jablka, konzumovateľnosť, skrátka každý elemntárny prvok skúsenosti s jablkom. Je to rovnaké ako v organickom svete. Každý pes má ňufák. Jeden je väčší, iný menší, jeden je vystúplejší iný menej, ale vždy to je charakteristický znak, ktorý sa rozmnožuje od psa k psovi a stále sa zachováva. Aj mém, ako charakteristické usporiadanie časti prostredia môže mať vždy trocha inú vnútornú štruktúru, ale vždy má charakteristické celkové usporiadanie.
Vyjadrime doteraz povedané v darwinovskom evolučnom mechanizme:
- Mém je informácia o prostredí, zapísaná v organizme ako časť modelu prostredia.
- Mém má schopnosť sebaopakovania — replikácia.
- Replikácia prebieha s rozličnou presnosťou — variabilita.
- Potomkovia pôvodného mému súťažia o obmedzené zdroje. Zdrojmi sú modely prostredia, napríklad mozgy — konkurencia.
- Tie mémy, ktoré sú schopné získavať zdroje rýchlejšie, či sa vo viacerých kópiách v jednom cykle replikovať, majú viac potomkov a postupne v populácii prevážia — selekcia, prírodný výber.
Prvé tri body sme si v predošlom výklade prebrali. O konkurencii a selekcii bude reč neskôr. Teraz si však skúsme rozobrať dôsledky darwinistickej evolučnej interpretácie mému.
- Prvý dôsledok je evolučná kontinuita. Mém sa neobjavil zrazu aktom nejakého zázraku, ale je tu ako etapa miliardy rokov trvajúceho vývoja. Mém je informácia, ktorá sa rozmnožuje z prostredia do organizmu. Tak tomu však bolo vždy, len sa to dialo geneticky, nie skúsenostne. Ak prežije jedna jediná baktéria v zhubnom prostredí antibiotika, pretože jej potomstvo do seba zreplikovalo informáciu o molekulách antibiotík do genetickej informácie, tak je to v princípe taká istá replikácia informácie, ako keď sa mláďa šimpanza naučí od svojej matky ako sa rozbíjajú orechy.
- Druhý dôsledok vyplýva z prvého. Pokiaľ chápeme mozog ako orgán modelovania prostredia, mém je elementom vnútornej organizácie tohoto modelu, inými slovami to čo považujeme za vedomie, myseľ, dušu je tvorené výhradne mémami. Navyše sa táto duša neobjavila až u človeka, ale sa postupne transformovala od štruktúr prvých organizmov a možno (ja tvrdím že určite) od samého vzniku tohoto Vesmíru. Štrukturovanosť a komplexnosť duše od prvých organizmov po dnes narastala tak, ako narastala štrukturovanosť a komplexnosť týchto organizmov.
- Tretí dôsledok poukazuje na to, že niet dôvodu hľadať na vysvetlenie akejkoľvek činnosti mozgu iné vysvetlenia a modely, ako také, ktoré vyplývajú z darwinistickej povahy elementov usporiadania mozgu — mémov. Každý psychický, či sociálny a napokon kultúrny jav musí byť vysvetliteľný pomocou evolučnej dynamiky a ekologických zákonitostí vyplývajúcich z replikačnej povahy mémov.
V ďalších kapitolách sa pokúsime postupovať presne v zmysle týchto dôsledkov a uvidíme, že striktne darwinistický prístup nám umožní jasne a nekompromisne uchopiť všetko, čo sa zdanlivo javí ako neuchopiteľné, záhadné a výhradne človečenské a to s pomocou malej skupiny základných predpokladov.
Subjektivita mému
Subjektivita mému
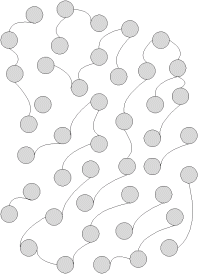
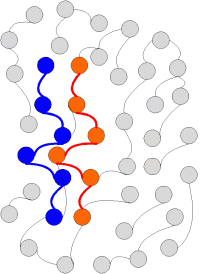
Pre ďalší výklad je dôležité pochopiť podstatnú vlastnosť mému, ktorá vyplýva z jeho replikácie. Replikáciu si môžeme schématicky znázorniť na nasledovných obrázkoch:
|

|
|
| obr. 1 | obr. 2 | obr. 3 |
- Na obrázku máme schématicky znázornenú časť bytia, skutočnosti, reality, prostredia, alebo ako si to nazveme. Krúžky znázorňujú prvky (entity) toho prostedia a čiarky ich vzájomné väzby. Napríklad je to časť biotopu, alebo je to kultúrne prostredie sapientov.
- Časť tejto reality znázornená na obrázku modrou farbou vystupuje ako jedna entita. Môže to byť bunka, gén na molekule DNA, jedinec, ale aj myšlienka, technický postup a každý prvok kultúry, teda mém.
- Podstatnou vlastnosťou takejto entity je, že sa rozmnožuje, replikuje ako ukazuje schématicky obrázok. Čo to znamená, že sa rozmnožuje? Znamená to tú skutočnosť, že sa zopakuje štruktúra entity v jej tvarovej stálosti, teda v pre ňu charakteristických väzbách a štruktúre. Takto sa stále zachováva druh organizmov, mémov, či iných entít. Mačka vždy vyzerá ako mačka, kryštál kremíka vždy vyzerá ako kryštál kremíka a trebárs slovo jablko vždy označuje nejakú našu skúsenosť s jablkom.
Tvarovú stálosť a jej replikovanosť považujem za základnú vlastnosť nášho Vesmíru. Diskusia o podstate tejto vlastnosti presahuje rámec tohoto cyklu. Tu stačí povedať, že aj mém je takýto spôsob zachovávania tvarovej stálosti skúsenosti, ktorá je v mozgoch reprezentovaná usporiadaním neuronálnej siete. Ako neskôr uvidíme, mém sa rozmnožuje (replikuje) nielen ako štruktúra nejakej časti prostredia do hlavy sapienta vo forme skúsenosti, ale aj medzi sapientami navzájom ako základný element sociálnej interakcie.
Darwinistický koncept replikácie nám však hovorí, že hoci sa zachováva tvarová stálosť (druh organizmov) existujú individuálne odchýlky, čiže variabilita. Replikácia teda nemusí byť presná, a to z dôvodov, ktoré sa súhrnne nazývajú náhoda. Inak povedané: presnosť replikácie prebieha s rôznou mierou pravdepodobnosti.
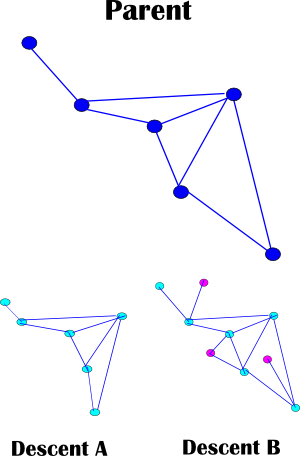
|
obr. 4 — Subjektivita

|
Túto skutočnosť si znázorníme na obrázku 4. Vidíme nejakú štruktúru rodiča (parent), ktorá má dvoch potomkov (descent) A a B. Základná štruktúra usporiadania rodiča sa zachovala, ale nie presne. Potomok A má niektoré väzby kratšie a potomok B má síce základnú štruktúru zachovanú, ale niektoré prvky v nej pribudli aj s väzbami.
Napríklad každý gén, uložený na molekule DNA je postupná štruktúra molekúl adenín, guanín, cytozín a tymín (AGCT). Štruktúra génu (napríklad ACCCTGGTCA) sa pri rozmnožení bunky zachováva. Potomkovia majú stále tú štruktúru ACCCTGGTCA. Niekedy však z rôznych dôvodov môže nastať zmena, napríklad jedno písmeno C je nahradené písmenom G — A G CCTGGTCA. Pre organizmus je dôležité, či proteín, ktorý sa pomocou takéhoto zápisu génu vyprodukuje plní v organizme danú funkciu. Ak áno, tak máme mutáciu, potomka génu, ktorý si zachoval svoju štruktúru. Ak sa pomocou takto pozmeneného génu produkuje nefunkčný, či dokonca organizmu škodlivý proteín, tak tento potomok génu nezachoval tvarovú stálosť a zanikne pod tlakom prírodného výberu.
Podobne si môžeme predstaviť našu štruktúru jablka, ktorá sa šíri ako skúsenosť medzi sapientami. Táto skúsenosť je síce individuálna, ale vždy zachováva základnú tvarovú skúsenosť s jablkom. Všetci čo toto čítajú mi rozumejú, ak už mali skúsenosť s jablkom. Súčasne mi rozumejú, pretože v rámci nášho jazykového okruhu je súčasťou individuálnej skúsenosti s jablkami aj od sapienta k sapientovi replikovaná štrúktura jazyka, v tomto prípade grafické symboly písmen spojené do slova jablko. Jablko ako skúsenosť a jablko ako pojem sa môžu v našom kultúrnom kontexte šíriť napriek tomu, že s ním má každý individuálnu slkúsenosť. Teda niečo sa zachováva. pričom sa to stále replikuje.
V tomto procese replikovania, kde sa niečo neustále zachované prenáša je podstatné nie odkiaľ sa replikuje, kam zreplikuje, ale to, čo sa pri replikácii zachovalo. Je jedno, či gén vyzerá individuálne tak, či onak, je jedno ako ja individuálne vnímam jablko a ako ho individuálne vníma čitateľ týchto riadkov, podstatné je to, že gén je tým génom ako jeho predok a jablko moje, či čitateľove je „jablkové“, čiže tiež má spoločného predka. Každá kópia replikovaného tvaru je iba okamihom v procese replikovania. Ak bolo pôvodné usporiadanie entít pred replikáciou nejaké, po replikácii sa mení v prospech usporiadania replikovaného tvaru.
Túto tendenciu k zachovaniu tvaru pri replikácii si zadefinujeme ako subjektivitu replikátora.
Budeme hovoriť, že mém, či gén chce zachovať svoju štruktúru, že je v jeho záujme byť rozmnožený v čo najviac kópiách, že uzatvára symbiózy s inými mémami, či génmi v záujme vlastného zachovania a podobne. Budeme tým myslieť to, že príroda uprednostňuje zachovanie tvaru vždy, keď je to ešte možné.
Doteraz sa pojem subjekt vždy prisudzoval človeku, ktorý v duchu kresťanskej tradície bol obdarený dušou, ktorá mohla subjektívne konať. Tento koncept sa vcelku hladko zreplikoval aj do racionalistickej vedy a prakticky nezmenený ostal vo filozofických konceptoch. Pri pokusoch o vysvetlenie čo to je subjektivita, však tieto koncepcie nemali rámec, z ktorého by vychádzali. Filozofie preto subjektivitu spravidla zahmlievajú a veda ponúka bezradne „záhadu vedomia“. Je až zarážajúce, ako mnohé myšlienkové koncepty, ktoré o sebe tvrdia, že skúmajú objektívne zákonitosti sveta súčasne pripúšťajú od objektivity nezávislý skúmajúci subjekt.
Darwinizmus vracia človeka späť do prírody. Z človeka nadaného subjektivitou robí to, čím v skutočnosti je — sapienta, ktorý je výslednicou minulých a súčasnych prírodných dejov, ktorý je nevydeliteľnou súčasťou prírody. A ak sa darwinizmus uplatňuje dôkladne, tak niet najmenšieho dôvodu, aby zachoval predstavu samostatne, od prírodných procesov nezávislej duše, či vedomia, niet dôvodov aby mu prisúdil subjektivitu. Úlohou darwinizmu je subjektivitu vysvetliť, pričom sa nemôže oprieť o doterajšie koncepcie, ktoré nepredpokladajú evolučné zákonitosti. Tak ako nechápe telesnosť sapienta ako jeho subjektivitu, ale ako dôsledok subjektivít evolučných procesov, tak musí to isté vykonať aj s jeho socialitou. Nikoho ani nenapadne tvrdiť, že človek má čiernu farbu pokožky, pretože sa mu to páči, ale preto, že to je výsledkom toho, aké má gény po predkoch, čiže to je dôsledkom subjektivity génov. Pri tom sa v minulosti stávalo, že keď sa belochom narodilo čierne dieťa, tak to bolo pripisované ich subjektivite — čarodejníctvu alebo hriechu. Súčasne však každý predpokladá, že má svoje myšlienky preto, lebo ich sám vymyslel, teda, že má subjektivitu. Súčasne však nevie vysvetliť, ako by telo, ktoré je výslednicou subjektivity génov stvorilo tú od doterajšieho evolučného vývoja nezávislú subjektivitu myslenia.
V teórii mémov sa pokúsim ukázať, ako sa tými istými procesmi, ktoré formujú naše telo formuje aj naše správanie a to čomu hovoríme myslenie a tiež ako sa úplne nezávisle od individuálity sapientov formuje ich kultúra, ba presnejšie: ako sa v záujme subjektivity mémov tvoria jednotliví sapienti tak, aby si mémy vytvárali vlastný ekosystém.
Od duše ku kultúre
Od duše ku kultúre
Ak
by sa mém replikoval iba z prostredia do organizmu, nebolo by potrebné
už ďalej o ničom hovoriť. Zaujímavé javy nastanú vtedy, keď sa mémy
začnú replikovať medzi jedincami.
Spoločenské organizmy majú oproti nespoločenským niekoľko výhod, najmä:
V skutočnosti je obsahom teórie mémov hľadanie ekologických javov v kultúrach, ich identifikácia a popis. Kým k tomu prikročíme, považujem za dôležité zdôrazniť dva aspekty veci.
Prvý dôležitý aspekt je, že takýto koncept teórie mémov je striktne darwinistický, pretože ukazuje, ako sa mémy postupne vyvíjajú od individuálnej skúsenosti potrebnej pre modelovanie prostredia, cez spoločenskú skúsenosť v sociálnej komunikácii po spoločenský mém, ktorý vytvára vlastný neorganický svet. Mém sa teda neobjavuje nejakým nekauzálnym činom, ale striktne darwinisticky, ako výsledok kauzálnej reťaze postupných adaptácií na prostredie.
Druhý aspekt veci je v tom, že sociálny mém môžeme predpokladať u iných sociálnych druhov, ktoré získavajú skúsenosť učením od jedincov svojej society. Takéto society majú teda tiež svoje kultúry.
Ako príklad môžeme uviesť našich najbližších príbuzných — primátov. U primátov boli pozorované rozdiely v správaní medzi societami, rozdiely, ktoré sú odovzdávané učením — replikovaním. Napríklad sú rozdiely v tom, ako rozbíjajú orechy jednotlivé tlupy šimpanzov, rovnako v iných metódach získavania potravy, tak isto napríklad v spoločných rituáloch. Ale nielen naši najbližší príbuzní. Boli pozorované podobné rozdiely v správaní napríklad jednotlivých kŕdloch vtákov, ale aj delfínov, či veľrýb.
Mémy a kultúra teda nie sú opäť v zmysle striktného darwinizmu ničím ľudsky výnimočným, vytvoria sa v podobných podmienkach vždy tak, ako sa vytvoria plutvy, či aerodynamický tvar u organizmov, ktoré žijú vo vode, alebo tvar krídla u organizmov, ktoré lietajú.
Spoločenské organizmy majú oproti nespoločenským niekoľko výhod, najmä:
- spoločná obrana pred predátormi,
- kolektívny lov,
- spoločná obrana teritória,
- zjednodušený pohlavný výber,
- a pre túto tému dôležitá možnosť odovzdávania výhodných skúseností.
Prirodzene, za tieto výhody sa platí, napríklad:
- náklady na prispôsobovanie sa ostatným,
- náklady na zložitejšie štruktúry správania sa (sociálne správanie), teda zložitejší model prostredia spojený s hladnejším mozgom,
- náklady na výchovu potomstva, ktoré sú spojené s učením potomkov, teda v predlžovaní obdobia neoténie (neoténia je obdobie od narodenia po reprodukovanie ďalších potomkov), alebo inak: vyššie rodičovské investície.
Pre
tento seriál je dôležité si uvedomiť, že individuálne skúsenosti, čiže
individuálne repliky mémov sa v prostredí spoločenských organizmov môžu
šíriť ďalej, od jedinca k jedincovi. Aby to bolo možné, musí byť mozog
na to pripravený, musí mať schopnosť napodobovať správanie sa druhých
jedincov v societe.
Túto
schopnosť majú mnohé druhy organizmov, hlavne cicavce, pravda v rôznej
miere. Schopnosť imitovať správanie iných je zrejme spojená s funkciou
tzv.
zrkadliacich neurónov
(
mirror neurons
),
ktoré boli objavené a potvrdené u primátov a vtákov. Zrkadliace neuróny
sú aktivované rovnako pri vlastnej činnosti ako pri pozorovaní činnosti
niekoho iného. Napríklad ak niekto dvíha predmet a ja to pozorujem,
zrkadliace neuróny v mojom mozgu sú rovnako aktivované, ako keď sám
dvíham nejaký predmet.
Z pohľadu teórie mémov ide o základný replikačný mechanizmus na replikovanie skúseností od jedinca k jedincovi.
Pre
takto replikované skúsenosti je potrebné vytvoriť v mozgu štruktúry, čo
je zrejme u cicavcov dôvodom pre vývoj čelných lalokov mozgu.
Neuronálna štruktúra mozgu je však asociatívna (k téme asociatívnosti sa
ešte vrátime), čo mimo iné značí aj to, že sa jej štruktúra vytvára
individuálnou históriou replikácií mémov do hlavy. Každý jedinec má inú
individuálnu históriu, a preto tie isté mémy odovzdávané od jedinca k
jedicovi vytvoria v mozgoch iné asociatívne štruktúry. Spoločensky
replikovaný mém má teda z darwinistického hľadiska vždy potomkov s
trocha inou štruktúrou, čím je zabezpečená diverzita replikácie. Napriek
diverzite, ostáva určitá základná usporiadanosť, inak by nebola možná
komunikácia v societe.. Povedané darwinisticky: v procese replikácie
mémov medzi jedincami prežijú iba také, ktoré je prijímajúci jedinec
schopný asociovať do vlastnej štruktúry, teda na replikáciu mémov pôsobí
prírodný výber.
Mémy,
ktoré sú schopné replikácie v rámci societ získavajú dôležitú
vlastnosť: stávajú sa nezávislé od jedincov, ktorí ich replikujú. Kým
replikácia vlastností prostredia do individuálnej skúsenosti závisela v
podstatnej miere na štruktúre jedinca, teda na jeho génmi determinovanej
stavbe, replikácia mémov medzi jedincami je od individuálnej stavby
jedincov nezávislá, a preto nadobúda význam vzájomných vzťahov medzi
mémami, teda význam
ekológie mémov
, o čom bude samozrejme reč v ďalších kapitolách.
Táto
nezávislosť nie je úplná, pretože je podmienená štruktúrou mozgov, a tá
je zasa podmienená druhovým genómom. Napriek tomu neskôr uvidíme, že
podobne ako v organickom svete, tvorivá sila prírodného výberu dokáže
prekonať mnohé obmedzenia a vytvorí také štruktúry mémov, ktoré umožnia
prežitie mémov, ktoré sú pre genómovú štruktúru replikujúcich jedincov
úplne neprirodzené.
Uvedené nám umožňuje vytvoriť si jednoznačnú definíciu kultúry:
Pod kultúrou budeme ďalej rozumieť ekosystém mémov, ktoré sú zdieľané určitou populáciou.
V skutočnosti je obsahom teórie mémov hľadanie ekologických javov v kultúrach, ich identifikácia a popis. Kým k tomu prikročíme, považujem za dôležité zdôrazniť dva aspekty veci.
Prvý dôležitý aspekt je, že takýto koncept teórie mémov je striktne darwinistický, pretože ukazuje, ako sa mémy postupne vyvíjajú od individuálnej skúsenosti potrebnej pre modelovanie prostredia, cez spoločenskú skúsenosť v sociálnej komunikácii po spoločenský mém, ktorý vytvára vlastný neorganický svet. Mém sa teda neobjavuje nejakým nekauzálnym činom, ale striktne darwinisticky, ako výsledok kauzálnej reťaze postupných adaptácií na prostredie.
Druhý aspekt veci je v tom, že sociálny mém môžeme predpokladať u iných sociálnych druhov, ktoré získavajú skúsenosť učením od jedincov svojej society. Takéto society majú teda tiež svoje kultúry.
Ako príklad môžeme uviesť našich najbližších príbuzných — primátov. U primátov boli pozorované rozdiely v správaní medzi societami, rozdiely, ktoré sú odovzdávané učením — replikovaním. Napríklad sú rozdiely v tom, ako rozbíjajú orechy jednotlivé tlupy šimpanzov, rovnako v iných metódach získavania potravy, tak isto napríklad v spoločných rituáloch. Ale nielen naši najbližší príbuzní. Boli pozorované podobné rozdiely v správaní napríklad jednotlivých kŕdloch vtákov, ale aj delfínov, či veľrýb.
Mémy a kultúra teda nie sú opäť v zmysle striktného darwinizmu ničím ľudsky výnimočným, vytvoria sa v podobných podmienkach vždy tak, ako sa vytvoria plutvy, či aerodynamický tvar u organizmov, ktoré žijú vo vode, alebo tvar krídla u organizmov, ktoré lietajú.
Základná úroveň kultúry
Základná úroveň kultúry
Je už dlhšie dokázané, že jazykové štruktúry máme vrodené. Štúdia [1] to v prípade slabík, ktoré sú vlastne základné jazykové tehličky, dokazuje. U novorodencov (vzorka priemerne 2,9 dňa stará) sa pri ich počúvaní rôznych slabík merala infračervená mapa ich mozgov. Ako autori uvádzajú, slabika blif je mozgami novorodencov zreteľne rozoznaná ľahšie ako slabika lbif, či bdif (vedel by som si predstaviť vhodnejšie príklady slabík, ale ďalej budem blif používať ako príklad). To značí, že určité slabiky sú pre všetky jazyky vrodené.
Tak toto u šimpanzov nenájdeme, Tí majú svoj jazyk iba v podobe nonverbálnych vrodených štruktúr gest a mimiky. A zrejme tie nepatrné génové rozdiely medzi šimpanzmi a sapientmi, ktoré nejako súvisia so stavbou mozgu, súvisia prevažne so stavbou vrodených jazykových štruktúr. Inak je zaujímavé, že podobné štruktúry boli zistené u vtákov, teda tiež organizmov, ktoré majú akustický jazyk. Zaujímavé by taktiež bolo robiť niečo podobné kytovcami a delfínmi.
Z pohľadu mémologického sú vrodené slabiky vrodenými mémami. Zrejme sa ich tvar, ich štruktúra, vyselektovala v konkurencii iných možných tvarov ako výhodnejšia. Skutočne blif sa aj mne ľahšie vysloví ako lbif. Zas na druhej strane bdif je v podstate s lbif rovnako náročné. Ale vývoj z nejakých dôvodov uprednostnil blif.
Pre každý mém je vyýhodné s mémom blif vytvoriť mémplex, pretože má väčšiu šancu sa rozšíriť do hláv ako ho vytvoriť s mémom lbif. Tu je vidno elementárnu ukážku tvorby mémosféry, kde určité mémy sú mozgami uprednostnené pred inými. Je to tá najzákladnejšia ekosystémová úroveň. Jazyk je spôsob na šírenie mémov, pretože pomocou jazyka sa už dokážu hromadne rozširovať v populácii sapientov.
Všimnime si podstatnú stránku veci, a to, že mém blif je vrodený. To značí, že jeho replikácia je zabezpečená genticky a jeho výsledná štruktúra je dôsledkom tlaku prírodného výberu na rôzne varianty u našich veľmi vzdialených predkov. Kedže je tento mém vrodený a daný evolúciou génov, je vlastne súčasťou znakov organizmu sapienta, teda súčasťou jeho fenotypu. Rovnako ako tvar hlavy, či počet končatín.
Definícia: α-mém je mém, ktorý bol vygenerovaný ako fenotypový znak biologickou evolúciou.Mozog obsahuje rad vrodených štruktúr pozostávajúcich z α-mémov.
Príkladom takejto štruktúry je napríklad tvar tváre, ktorý máme vrodený a rozoznávajú ho už novorodenci [2]. Ale aj vrodený modul jazyka. Mozog je asociatívny systém, to značí, že novú informáciu skúsi podľa podobnosti jej štruktúry nájsť vo svojich štruktúrach a ak také nájde, vytvorí iba tie nové časti informácie, ktoré ešte v sebe nemá a tie priradí k informáciám stávajúcim. Znázornime si to graficky:
| Obr.1: stav v mozgu pred asociáciou mémov | Obr.2: stav v mozgu po asociácii mémov |

|

|
Aby sa dokázal nejaký n-mémplex usídliť v mozgu, tak si to výrazne zjednoduší ak obsahuje α-mém. Mozog dokázateľne uprednostňuje informácie, ktoré už nejako pozná. Na ich začlenenie potrebuje vynaložiť menej energie ako na úplne nové informácie. To je známe aj z pedagogiky, kde sa hovorí to isté, len inak: učiť treba v známych kontextoch. Na obrázku 1 vidíme nový mémplex, n-mémplex, ako sa snaží vreplikovať do mozgu. Mozog v ňom rozoznal α-mém. Zvyšok mémov n-mémplexu sa musí naučiť, teda musí pre ne vytvoriť neuronálnu štruktúru. Pre α-mém je to ovšem výhodné, pretože už existuje v dvoch kópiách, teda v dvoch kontextoch, v dvoch mémplexoch. Jeho šanca na prežitie stúpla.
Ak je blif vrodený α-mém, tak je pre každý iný mém výhodné s týmto mémom vytvoriť mémplex. Dá sa dobre predstaviť, že v evolučnej minulosti si naši predkovia nejaké slabiky, fonémy odovzdávali ako signály k určitým situáciám. Mozgy, ktoré dokázali takéto fonémy ľahšie tvoriť mali selekčnú výhodu, a preto postupne v populácii prevážili. Vytvorili jazykový f-mémplex, ako nástroj jednoduchej komunikácie.
Od tohoto momentu ovšem vývoj mémlexov, obsahujúcich α-mémy prestáva byť jednoznačne v réžii biologickej evolúcie. Pre mozog je podstatný α-mém, ktorý sa v biologickej evolúcii vyvinul. Ale mémy, ktoré sa s ním spoja už podliehajú evolúcii nebiologickej. Či sa blif spojí so situáciou A, alebo B záleží od okolností, v ktorých sa sapient nachádza. Teda obsah n-mémplexu závisí od ekosystému sociálnych interakcií medzi sapientami. Napríklad α-mém blif v spojení s α-mémom í mohol značiť pôvodne „som spokojný“, ale aj trebárs „som pokojný“ a všeličo podobné. To, aký význam spojenie týchto α-mémov nabralo, závisí od spojení s inými mémami, od sledu udalostí, v ktorých sa to stalo a od radu iných okolností. Typická evolučná situácia. Veľa variánt a z nich selekcia. Výsledok je element kultúry, teda význam. Význam je spojenie α-mémov s mémami, ktoré sa presadili prírodným výberom v prostredí sociálnej interakcie.
Takéto spojenie, teda takýto n-mémplex sa učením prenáša z generácie na generáciu. Pre všetky mémy je výhodné, ak sa im podarí s ním vytvoriť mémplex. A tak v mozgoch vzniká hierarchická štruktúra mémplexov, ktorú vedeckým jazykom popisuje aj [2]. Obsah a štruktúra mémplexov významu má svoj vlastný život. Každý sapient, ktorý sa narodí, nie je determinovaný len vlastnou genetickou evolučnou minulosťou, ale aj prostredím, ktoré si rovnako nevybral. Súčasťou toho prostredia sú ovšem aj kultúrne štruktúry, teda ekosystém mémov. Tie postupne prechádzajú z predkov na potomkov a dotvárajú štruktúru mozgu. Ako ten, ktorý mozog napokon dopadne, závisí od ekosystémových zákonitostí toho prostredia, nie od samotného sapienta. Napokon tak je to aj s jeho génmi. To ako ktorý variant génu dopadne v danom prostredí nezávisí od neho, ale od prostredia.
Vrodené jazykové štruktúry sa takto stávajú základným nástrojom, pomocou ktorého si mémy budujú svoj ekosystém. Pretože každý sapient sa nemôže vyhnúť tomu, aby bol donútený jazyk používať, je súčasne donútený replikovať ekosystém mémov, na vývoj ktorého má rovnako nepatrný vplyv, ako má gén na vývoj biosféry.
- Gómez, D. M., Berent, I., Benavides-Varela, S., Bion, R. a H., Cattarossi, L., Nespor, M., & Mehler, J. (2014). Language universals at birth. Proceedings of the National Academy of Sciences of the United States of America, 111(16), 5837–41. doi:10.1073/pnas.1318261111, http://www.sissa.it/cns/Articles/Gomezetal2014pnas.pdf
- Zhen, Z., Fang, H., & Liu, J. (2013). The Hierarchical Brain Network for Face Recognition. PLoS ONE, 8(3), e59886. doi:10.1371/journal.pone.0059886
Domestikácia sapienta
Domestikácia sapienta
Prakticky od vzniku modernej biológie v 18. storočí sa v kolísavej intenzite vynára v biológii otázka podobnosti sapienta s ním domestikovanými druhmi organizmov, najmä cicavcov. Fenotypová podobnosť je taká zarážajúca, že je veľmi ťažké argumentovať proti predstave sapienta ako jediného domestikovaného druhu opice.
Pokiaľ túto predstavu akceptujeme, vynára sa celý rad otázok, najmä otázky ako k domestikácii došlo a prečo. V doterajších teóriách sa predpokladá, že sapienta domestikoval človek, teda, že sa domestikoval sám — tzv. teória autodomestikácie. Implicitným predpokladom teórie autodomestikácie je vyčlenenosť sapienta z prírodných zákonitostí. Predsa Človek domestikoval psa, sviňu, či sliepku. Niet známeho organizmu, ktorý by robil to isté sapientovi — len Človek. Ten sa však rozhodne nedomestikoval zámerne, ale iba ako vedľajší efekt iných svojich prejavov. Nepoznáme iné vedľajšie životné prajavy sapientov iba jeho kultúru. Pokiaľ chápeme kultúru antropocentricky, ako produkt vedomej, zámernej činnosti Človeka, dostávame sa do situácie, kedy predpokladáme, že Človek vedomou činnosťou domestikuje sám seba, čo určite znie prinajmenšom trocha nepresvedčivo.
Pokiaľ však domestikáciu chápeme ako vedľajší efekt vedomých činností jednotlivých sapientov, ako ich nevedomú výslednicu, dostávame sa do do situácie, ktorá je v rozpore s predstavou autodomestikácie. Ak domestikačný tlak vytvára niečo, systém, výslednica, tak už vôbec nemožno hovoriť o nadradenosti a nezávislosti na prírodných procesoch, ale na ich závislosti. V takom prípade však je domestikátorom ten systém s jeho vlastnými potrebami a zákonitosťami, teda s jemu vlastnou intencionalitou. Som toho názoru, že teória mémov je jediná schopná túto intencionalitu popísať, a tým prispieť k pochopeniu autodomestikácie.
V ďalších kapitolách sa pokúsim aspoň zhruba popísať otázky spojené s domestikáciou obecne a s domestikáciou sapienta špeciálne z pohľadu teórie mémov.
Domestikácia
Domestikácia
Pojem domestikácie je prakticky nemožné jednoznačne vymedziť. Je šváb, mucha domová, myš domová či potkan domestikovaný organizmus? A ak nie, čím sa líši trebárs od včely medonosnej, alebo laboratórnej myši, či laboratórneho potkana? Alebo skúsme z inej strany. Je austrálsky zdivený pes Dingo nedomestikovaný, keď stále vykazuje znaky domáceho psa? Sú mačky a psy, ktoré sa potulujú v okolí sapientích sídiel a živia sa ich odpadkami nedomestikované? V čom sú iné ako tatranské medvede, navyknuté vykrádať kontajnery?
Aké kritérium si pre domestikovanosť vybrať? Všetci, čo sa hlbšie zaoberajú domestikáciou ako biologickým procesom veľmi skoro narazia na nepríjemný problém antropocentrizácie a antropomorfizácie kritérií domestikácie. Dlho sa napríklad chápalo, že domestikované organizmy boli domestikované Človekom pre jeho potreby, teda ako vedomý a uvedomelý akt, s cieľom získať hospodársky úžitok.
Ibaže bližší pohľad na vec ukazuje obtiaže tejto koncepcie. Mnohé o tom hovorí a píše prof. Stanislav Komárek, napríklad v [1]. Niektoré druhy organizmov sa k našim predkom dostali akosi sami od seba. Príkladom môže byť mačka domáca, ktorá sa objavila pri sapientovi od okamihu uskladňovania obilia spolu s myšami v sýpkach. Takýmto organizmom hovoríme sinantropické, teda žijúce v blízkosti sapienta. Sem patrí aj spomínaný šváb, či mucha domová a keby sa veci ponechal voľný priebeh, zrejme by sa sinantropom stal aj tatranský medveď. Od sinantropizácie k ešte bližšiemu súžitiu a nahradeniu prírodného výberu hospodárskou selekciou výhodných jedincov je predstaviteľný malý krok. Nie vždy si to však vieme predstaviť. Napríklad ako Komárek v [1] uvádza:
Domestikace velkých zvířat typu koně, či velbloudi velice připomíná problémy se začátkem metalurgie. Ono se to tak vykládá, jak nějací ti neolitici, či chalkolitici, či co to bylo si jednoho dne obložili ohniště řekněme chasiteritem a vytavili si ten cín. No skuste si to někdy udělat, co se vytaví. Nápad skoušet něco, co by k metalurgii mohlo vést, je neobyčejně odlehlý od všeho, co běžně lidé podnikají a zůstává to mystériem sui generis. Právě tak třeba domestikace koní. Vždycky jsou pro toto zajímavé takové ty neodomestikační pokusy, třeba s koňmi Przewalského, kdy firma Hagenbeck pomocí bratrů Asamovových asi v roce 1910 nechávala toto atraktivní a nové zvíře chytat a dopravovat do Evropy. Jak se to dělalo… Inu to divoké stádo se jaksi obeznalo, kde je, potom se v stepi umístnili skupiny doma osedlaných domácích koní v takovém hodně velikém kruhu a začala štvanice. Ti divocí koně se hnali po obvodu té velekružnice. Pronásledovatelé si vždycky přesedali na nové čerstvé koně, a tak po čtyřech hodinách začala najmladší, několidenní hříbata umdlévat. Bylo možno pak zastřelit vedoucího hřebce a ta hříbata pochytat, načež se podložila domácím klisnám, vzhledem k tomu, že jsa krmena mlékem koz nebo ovcí pocházela na těžké průjmy
Toto samozřejmě nechává vzejít otázku: jakým spůsebem se opěšalí obyvatelé střední Ázie prvních koní zmocnili a jakým spôsobem začali ten chov? Je to samozřejmě tak trochu jako když baron Prášil v té své povídce sebe sama vytahuje za vlasy z bažiny… I kůň se zlomenou nohou je nález špatný, vzhledem k tomu, že ty zlomeniny dlouhých kostí jim srůstají neobyčejně těžko, prakticky vůbec. Nález čerstvě nalezeného hříběte, od něhož odženeme klisnu, zase dává obtíž, jakýmže mlékem ho vlastně vychovat.
Podobne s nadsázkou Marco Stella v [2] ironizuje predstavu cielenej domestikácie:
…za tím stojí představa člověka jako pána tvorstva, toho prvního supergeniálního zemědělce, kterej příde a teďka si řekne: tak copak to tady roste? Výborně, tak z týhle hnusný, hořký a jedovatý věci já udělám salát a támhle z tý vobludy, která mě včera pokopala a pokousala udělám velblouda…
Taktiež je napríklad veľmi obtiažne si predstaviť ako mohol niekto v tejto rastline vidieť budúcu kapustu, kareláb, karfiól, brokolicu, horčicu, či kvaku. Šľachtiteľstvo, ako ho dnes poznáme, teda kríženie a selekcia želaných typov, je známe až od 19. storočia. Do tej doby zrejme domestikátor uprednostňoval z prirodzene produkovanej diverzity jedincov tie typy, ktoré mu vyhovovali a tie ostatné ignoroval alebo likvidoval [1].
Podobných príkladov je v uvedeých zdrojoch viacej. Predstava uvedomelej selekcie ako domestikačného aktu je skrátka ťažko udržateľná. Zdá sa, že oveľa udržatelnejšia je predstava postupného zbližovania dvoch organizmov: domestikátora a domestikanta, teda postupná sinantropizácia domestikovaného druhu.
Z mnohých štúdií (napr. [3]) ďalej vyplýva, že domestikácia rozhodne nie je doménou sapienta. Aj u mravcov prebieha domestikácia niektorých iných druhov organizmov a zrejme podobným spôsobom, podľa autorov na základe podobnosti domestikovaného organizmu s juvenilnou formou domestikátora. Naši predkovia zrejme uprednostňovali z variability domestikovaných jedincov tie, ktoré viac pripomínali mláďa sapienta. Podobne aj mravce si z rôznych foriem hmyzu vybrali očividne tú, ktorá dotykovo a feromonálne viac pripomína larvy mravcov.
Na domestikáciu sa môžeme pozerať aj otázkou, kto vlastne má z domestikácie prospech? Antropocentricky sa obvykle predpokladá, že samozrejme domestikátor. Pravdou však je, že každý domestikovaný druh sa oproti svojej divej forme rozšíril do nebývalých počtov a teritórií. Často vytlačil z ich prirodzených biotopov pôvodné divé formy. Iste, odmenou za toto rozšírenie je individuálny osud v prospech domestikátora, čím sa to však vymyká z iných ekologických vzťahov v prírode, kde každý niečo obetuje v prospech iného a sám ťaží z obetí iných? Počíta sa reprodukčný úspech. Prospech z domestikácie je určite obojstranný. Bez hospodárskych plodín a zvierat by sotva bol možný taký reprodukčný úspech druhu Homo sapiens a bez Homo sapiens by nebol možný enormný reprodukčný úspech domestikantov.
Zhrňme si dosial preukázané:
- Domestikácia neprebehla ako uvedomelý plánovaný akt.
- Domestikácia nie je výlučnou doménou druhu Homo sapiens.
- Domestikácia je obojstranne prospešný stav pre domestikovaného a aj pre domestikujúceho.
- Domestikácia prebehla v mnohých prípadoch pravdepodobne formou sinantropizácie, nie je preto možné stanoviť, kto koho domestikoval
Z pohľadu biológa je domestikácia teoretický problém. Vidno to naapríklad v rôznych definíciách domestikácie u rôznych autorov. Povedzme taká ešte čiastočne antropocentrická definícia autority na správanie domestikovaných zvierat Pricea [4] , kde domestikácia:
… je proces, v ktorom sa populácia zvierat adaptuje na človeka a na podmienky prostredia, v ktorom je zajatá, genetickými zmenami pretrvávajúcimi generácie…
Táto definícia zjavne nezahŕňa súžitie mravcov s ich domestikantmi a sapient tu hrá ešte stále určitú centrálnu úlohu. Komárek v [1] definuje veľmi opatrne domestikáciu ako:
… mutuální proces interakce dvou živých organizmů…
V podstate každá symbióza sa dá definovať takto a skutočne — v čom je principiálny rozdiel medzi spolužitím sasanky s krabom a sapienta s prasaťom? V oboch prípadoch sa to bez mutuálnych zmien u oboch symbiontov nezaobíde, či už sú to zmeny genetické, alebo zmeny v spôsobe života kultúrne.
Niekde medzi týmito dvoma definíciami je svojou podstatou definícia Kleisnera a Stellu [3]:
Domestikácia je evolučný proces genetických a ontogenetických adaptácií organizmov na podmienky kultúry. Súčasne domestikované zvieratá tvarujú, udržiavajú a rozširujú kultúrne prostredie.
Ak pominieme, že sa zo všetkých foriem domestikácie touto definíciou diskvalifikujú rastliny, autori chápu ako selektor prírodného výberu v procese domestikácie kultúru, hoci ju nikde bližšie nešpecifikujú. Vec však je tak očividná, že je pre všetkých, čo sa domestikáciou zaoberajú implicitná, aj keď nevysvetlená. Pokiaľ však chápeme kultúru ako ekosystém mémov, zákonitosti tvoriace selektory prírodného výberu v procesoch domestikácie treba hľadať práve v ekologických zákonitostiach mémov.
Literatúra a odkazy:
- Komárek, S. (2009). Vepři, naši bratři — domestikace jako fenomén. Audioprednáška http://bio.natur.cuni.cz/~flegr/ctvrtky/historie.php?skrok=2008/2009
- Stella, M., & Havlíček, J. (2009). Skončila lidská evoluce? Autodomestikace vs. prostředí evoluční adaptovanosti. Audioprednáška http://bio.natur.cuni.cz/~flegr/ctvrtky/historie.php?skrok=2008/2009
- Kleisner, K., & Stella, M. (2009). Monsters we met , monsters we made : On the parallel emergence of phenotypic similarity under domestication, 37. http://web.natur.cuni.cz/filosof/images/stories/kleisner/5Kleisner_Stella.pdf
- Price, E. O. (1999). Behavioral development in animals undergoing domestication. Applied Animal Behaviour Science, 65(3), 245–271. doi:10.1016/S0168-1591(99)00087-8
Fenotyp domestikanta
Fenotyp domestikanta
Domestikované organizmy sa vyznačujú určitými spoločnými znakmi. Dokonca, pokiaľ sa budeme pohybovať vo vývojovej vetve cicavcov, dá sa hovoriť o fenotype domestikanta (napríklad v [1]), teda súbore morfologických a behaviorálnych znakov, ktoré sú spoločné pre domestikované zvieratá. Najprv si uvedieme niektoré z takýchto znakov hlavne z pohľadu domestikácie sapienta.
Morfologické znaky
Veľkosť tela
Oproti divej forme veľká variabilita vo veľkosti tela [1]. V pôvodnom ekosystéme divej formy je veľkosť tela pod silným selekčným tlakom. V domestikovanej forme je pod tlakom domestikátora. Podľa [1] je zaujímavé, že u viacerých druhov veľkých cicavcov je na začiatku domestikácie tlak na menšie veľkosti tela, zrejme z dôvodu ľahšej manipulovateľnosti. V neskoršej fáze býva tlak na veľkosť z dôvodu úžitkovosti.
Alometria
Teda vzájomný pomer rôznych častí tela. Podľa [1] je u domestikantov oproti divým formám častý výskyt obrích, či trpazličích foriem, vzájomný pomer častí je často posunutý do takých disproporcií, ktoré by v prirodzenom prostredí neumožnili prežitie. Kráke nôžky a dlhé telo jazvečíka, či trpazličie formy čivavy rozhodne nie sú vhodné pre prežitie v prírode.
Alometrický rast často zasahuje orgány, mozog a iné časti tela, čo má destabilizačný účinok na fyziológiu a správanie domestikanta (podľa [3]).
Podľa [1] je v procese domestikácie typická vysoká variabilita a fázová zmena veľkosti tela. Počiatok domestikácie obvykle sprevádza zmenšenie veľkosti tela z dôvodu ľahšej manipulovateľnosti s domestikantom. Nskôr naopak nasleduje zväčšenie z dôvodu vyššej úžitkovosti.
Kostra
Modifikácie kostry patria medzi záasadné symptómy domestikácie Často môžeme vidieť zmeny v počte stavcov, najmä chvostových stavcov pričom u niektorých druhov sa mení ich vzájomná poloha. Napríklad chvosty ošípaných, a tiež niektorých psov a mačiek, sa tvarovali do špirály [1]. V tejto súvislosti treba spomenúť slávne Beljajevove líšky, o ktorých ešte bude reč. Beljajev domestikoval sibírsku líšku a u nej sa tiež objavili niektoré formy s krútenými chvostíkmi [3].
Tieto kostrové zmeny sú takmer vždy spojené s úpravami tvaru hrudného koša a skrátením nôh. Všeobecne platí, že kosti domácich foriem sú obvykle širšie v priemere než u volne žijúcich príbuzných. Sú však krehkejšie a oveľa "nedbalejšie" spojené. Opäť sa dá povedať, že tieto zmeny by boli v pôvodnom prostredí prírodným výberom negatívne selektované.
Lebka
Kľúčovým znakom domestikácie cicavcov je skrátenie tvárovej časti lebky, ktorá je výstižne v nemeckej literatúre označované ako „Vermopsung“ (zmopslíkovnie) lebky. Vznikne tak, že sa relatívne zväčší temenná /parietálna) a spodinná (occipitálna) časť lebky. Príkladom nech je porovnanie lebiek vlka a buldoga aloebo svine divej a svine domácej:
 vlk |
 buldok |
 sviňa divá |
 sviňa domáca |
Zmopslíkovatenie lebky je podľa [1] a [2] kľúčové pre domestikáciu hlavne cicavcov, pretože takýto tvar lebky pripomína mláďacie rysy, čo zrejme útočí na rodičovské vrodené psychické moduly.
Časté bývajú aj zmeny chrupu, napríklad zmenšenie očných zubov u domácej svine.
Mozog
Podľa [1] je zmenšenie mozgu domestikanta oproti divej forme snáď najvýznamnejším a najspoľahlivejším znakom domestikácie. Príklady: pes 30%, sviňa domáca 24%, kôň 19%, ovca 24%… Redukcia mozgu sa pohybuje od 0% — 33% podľa [7] a postihuje najmä senzorické oblasti a cerebrum (telencephalon).
Povrch tela a pigmentácia
Pokrývka a pigmentácia tela býva u divých foriem pod silným tlakom prírodného výberu, pretože často plní ochrannú funkciu ([1], [5], [6]) a sebaprezentačnú funkciu ([1], [4]). U domestikovaných foriem často vidíme úplnú stratu ochrannej funkcie pigmentácie a jej vysokú variabilitu. Napríklad divá sviňa má jeden druh ochranného sfarbenia (čierny), kdežto u domácej svine vidíme širokú škálu farieb od bielej, cez ružovú, nahnedlú az po čiernu. Vyskytujú sa aj rôzne strakaté sfarbenia, s nepravidelným rozložením škvŕn [1].
Oproti divým formám je vysoká variabilita aj v ochlpení ([1], [5], [6]), čo je opäť umožnené tým, že prestáva pôsobiť unifikačný tlak prírodného výberu, vyplývajúci z pre konkrétny druh daného ekosystému. Rozloženie, hustota a tvar jednotlivých chlpov silne variujú [1].
Behaviorálne znaky
Z filozofického predpokladu nedeliteľnosti duše a tela a z evidencie znakovej polyfunkčnosti mnohých génov organizmov môžeme predpokladať, že domestikácia má vplyv nielen na vzhľad organizmov ale aj na ich správanie.
Nižšia miera agresivity
Dokázal to D. K. Belyaev, keď v 50-tych rokoch 20. storočia začal v Novosibirskom Inštitúte cytológie a genetiky dnes už legendárny experiment s domestikáciou sibírskych líšok [3]. Vychádzal z toho, že prvý krok akejkoľvek domestikácie musí byť selekcia na znášanlivosť domestikanta voči domestikátorovi. Ide teda o selekciu rysov správania, nie fyzických znakov. Nečakaným vedľajším výsledkom chovu boli typické vyššie uvedené domestikačné zmeny morfologické a fyziologické, napr. juvenilné rysy, modré oči, zmeny farby srste, pokrútené chvostíky a podobne.

|


|
|
Divá forma sibírskej striebornej líšky |
Domestikované formy sibírskej striebornej líšky |
Nižšia miera vnútrodruhovej agresivity je jednýmn z hlavných behaviorálnych znakov domestikantov [6].
Predĺženie neotenického správania.
So sníženou mierou agresivity súvisí zrejme aj neotenické správanie aj v pohlavnej dospelosti. Podľa [6] tlak na nižšiu mieru agresivity v konečnom dôsledku selektuje neotenické správanie, kedže mláďatá sú zákonite menej agresívne.
Ukazuje sa teda, že pravdepodobne fyzická užitkovosť zvierat nebola primárnym selekčným kritériom našich prapredkov, ale že si naprílad brali menej agresívne vlčie mláďatá ako hračku, alebo náhražku za vlastné mláďatá. Ostatne podobne to asi vo viacerých prípadoch robia v opačnom garde aj vlci s našimi mláďatami (tzv. vlčie deti) [2].
Neotenické správanie je spojené s jedným dôležitým rysom správania, a to s ponechaním riešenia rôznych životných situácií na svojich rodičov, na ich starostlivosť. Ilustratívne to preukazujú pokusy Adama Miklosiho z Eötvös Loránd University:
Vidíme dva pokusy v ktorých sa vlk chovaný od narodenia v zajatí správa ďaleko autonómnejšie ako pes, ktorý sa pri rozhodovaní spolieha na svojho pána. V prvom pokuse sa vlk rozhodne podľa vlastného čuchu správne pre potravu ukrytú vo vedierku, kým pes sa spoľahne na gesto svojho pána bez ohľadu na to, čo mu káže čuch. V druhom pokuse po predbežnom tréningu sa vlk snaží vlastnými silami získať potravu z klietky, kým pes to po chvíli vzdá a obráti sa na pána pre pomoc.
Predĺženie neoténie (často až do smrti) je veľmi výhodné pre domestikátora, pretože uľahčuje manipuláciu s domestikantom. Je však súčasne aj veľmi výhodné pre domestikanta, pretože mu to umožňuje vyriešiť životné situácie, na ktoré nie je prírodným výberom dizajnovaný jeho divoký predok, čo mu umožňuje prežiť aj v prostrediach, v ktorých by jeho predok neprežil.
Zvýšená sexuálna apetencia
Podľa [6] majú mnohé domestikované druhy zvýšenú sexuálnua apetenciu. To sa prejavuje často zvýšením počtu páriacich období. Vlk má napríklad jedno páriace obdobie v zime, kdežto pes má dve. Rovnako je tu častá aj nesezónnosť párenia.
Ďalším rysom zvýšenej sexuálnej apetencie je znížená vyberavosť partnera [6], spojená s nižším rozlišovaním partnera vôbec. Niektoré druhy sú schopné sa páriť aj s maketami.
Zvýšená apetencia potravná
Viaceré domestikované druhy majú zmenené stravné správanie ([5], [8]). To sa prejavuje neraz zmenou skladby potravy. Pes, sviňa domáca alebo mačka zožerú širšie spektrum potravín ako ich divokí príbuzní. Taktiež sa domestikácia prejavuje v prehnanej žravosti, čo je napríklad u svine domácej alebo domáceho vtáctva domestikátorovou selekciou podporované. Vyššia žravosť je ovšem pozorovaná aj u iných domestikovaných druhov, kde to nie v záujme domestikátora, napríklad u psa.
Patológia správania
U domestikovaných zvierat badáme rôzne formy patologického správania ([5], [8], [9]). Príkladom môžu byť rôzne duševné poruchy psov, napríklad naháňanie si vlastného chvosta, nečakané a neodôvodnené útoky. Rôzne formy stresového správania vidíme u klietkového chovu hydiny, či prasiat.
Iné zmeny správania
- Znížená lokomotorická aktivita
- Zmeny inštinktivních prvkov správania
- Znížená senzorická aktivita (Verarmung der Merkwelt)
Vysvetlenie medzidruhového fenotypu domestikanta
Vysvetlenie zvláštneho javu, pri ktorom sa určitým typom selekčného tlaku vyskytujú u rôznych druhov rovnaké alebo veľmi podobné znaky som nenašiel. Určité indície sa dajú nájsť v [6], kde sa vysvetlenie hľadá v samotných podmienkach domestikácie teda vo forme selekčného tlaku. To však nevysvetľuje fakt, že genotyp rôznych druhov musí byť na také zmeny pripravený, aby ich mohol vyjadriť vo fenotype.
Literatúra a odkazy:
- Kleisner, K., & Stella, M. (2009). Monsters we met, monsters we made : On the parallel emergence of phenotypic similarity under domestication, 37. Journal of Memetics (preklady článkov)http://web.natur.cuni.cz/filosof/images/stories/kleisner/5Kleisner_Stella.pdf
- Komárek, S. (2012). Ochlupení bližní: Zvířata v kulturních kontextech (p. 262). Axademia.
- Belyaev, D. (1979). Destabilizing selection as a factor in domestication. Journal of Heredity. http://jhered.oxfordjournals.org/content/70/5/301.short
- Portmann, A. (1960). Neue Wege der Biologie. München, R. Piper [1960].
- Stella, M. (2006). Marco Stella Teorie autodomestikace v německé antropologii a biologii do roku 1945. Kontext, důsledky, zastánci. 2006. http://www.fhs.cuni.cz/etologie/index.php/en/downloads/doc_details/5-teorie-autodomestikace-v-nemecke-antropologii-a-biologii-do-roku-1945-kontext-dusledky-zastanci
- Price, E. O. (1999). Behavioral development in animals undergoing domestication. Applied Animal Behaviour Science, 65(3), 245–271. doi:10.1016/S0168-1591(99)00087-8
- Guay, P.-J., & Iwaniuk, A. N. (2008). Captive Breeding Reduces Brain Volume in Waterfowl (Anseriformes). The Condor, 110(2), 276–284. doi:10.1525/cond.2008.8424
- Stella, M., & Havlíček, J. (2009). Skončila lidská evoluce? Autodomestikace vs. prostředí evoluční adaptovanosti. http://bio.natur.cuni.cz/~flegr/ctvrtky/historie.php?skrok=2008/2009
- Moberg, G. (1987). A model for assessing the impact of behavioral stress on domestic animals. Journal of animal science, 1228–1235.
Homo sapiens domesticus
Homo sapiens domesticus
Mnohé z morfologických a behaviorálnych znakov fenotypu domestikanta má aj druh, ktorý sám seba nazval Homo sapiens sapiens. Pri posudzovaní morfologických, fyziologických, či behaviorálnych znakov, ktoré by mohli byť vyvolané domestikačným tlakom narážame na niekoľko špecifických ťažkostí:
- Chýba divá forma, ku ktorej by sme mohli náš druh prirovnať podobne ako napríklad psa k vlkovi. Všetci sapienti na svete vykazujú domestikačné znaky. Hoci by sa tento fakt mohol zdať ako diskriminujúci predstavu sapienta ako domestikovaného druhu, množstvo paleontologických nálezov predchodcu nášho druhu, teda Homo erectus, taktiež množstvo nálezov poddruhu Homo sapiens neanderthalensis a v neposlednom rade neolitické nálezy nášho druhu do značnej miery podporujú predstavu domestikácie Homo sapiens sapiens. Súčasne treba povesať, že domestikácia je proces priebežný a nie konečný. Rôzne rozdiely možno teda pozorovať aj voči prírodným národom, kde predpokladám menšiu mieru domestikácie ako ju pozorujem u národov civilizovaných. Takisto možno aj v rámci jednotlivých populácií, či rás pomocou historických a archeologických záznamov pozorovať určitý vývoj (viď odkazy).
- Sapient je mimoriadne evolučne úspešný druh. Podobne ako sinice, či baktérie osídlil všetky kontinenty planéty. Z toho vyplýva záver, že treba odlíšiť vplyv na vnútrodruhové zmeny daný tlakom lokálnych prírodných podmienok od tých, ktoré sú vyvolané tlakom domestikácie. Napríklad rasové zmeny sú zrejme do veľkej miery spôsobené štandardným procesom tlaku prírodného výberu na oddelené populácie v iných klimatických a geologických podmienkach.
- Problém selekčných potrieb domestikátora. Vzhľadom na to, že na rozdiel od domestikácie organizmov sapientom vieme pomerne dobre identifikovať selekčné potreby domestikátora, u domestikácie samotného sapienta narážame na ťažkosti, vyplývajúce z teoretickej nejednoznačnosti doterajšej predstavy autodomestikácie. Vzhľadom na to, že táto práca považuje za domestikátora darwinistický ekosystém mémov, je táto ťažkosť podľa mojej mienky prekonateľná hľadaním evolučných zákonitostí v mémosfére planéty.
V ďalších kapitolách sa pokúsim predložiť hlavné znaky, ktoré môžu byť dôsledkom domestikácie sapienta. Tieto znaky nás oprávňujú chápať náš poddruh druhu Homo sapiens ako poddruh, ktorého výstižný názov by mohol byť Homo sapiens domesticus.
Literatúra a odkazy:
- Stella, M. (2006). Marco Stella Teorie autodomestikace v německé antropologii a biologii do roku 1945. Kontext, důsledky, zastánci. 2006. http://www.fhs.cuni.cz/etologie/index.php/en/downloads/doc_details/5-teorie-autodomestikace-v-nemecke-antropologii-a-biologii-do-roku-1945-kontext-dusledky-zastanci
- Stella, M., & Havlíček, J. (2009). Skončila lidská evoluce? Autodomestikace vs. prostředí evoluční adaptovanosti. http://bio.natur.cuni.cz/~flegr/ctvrtky/historie.php?skrok=2008/2009
-
Leach, H. M. (2003). Human Domestication Reconsidered. Current Anthropology, 44(3), 349–368. doi:10.1086/368119