Obsahom tejto knihy budú rôzne články a úvahy k práci na univerzálnej teórii mysle a kultúry človeka
Poznámky k mémológii
Poznámky k mémológii
Summa memologiae
Summa memologiae
Táto esej pôvodne vznikla ako mailová odpoveď na mailovú otázku počas vianočných sviatkov, čo sa nesporne odrazilo v jej uvolnenom štýle. Na výzvu adresáta ju napokon publikujem tak ako bola pôvodne formulovaná len s minimálnymi úpravami.
Aby sme na toto mohli nájsť odpoveď je dôležité vedieť z akého backgroundu budeme vychádzať. Môj je mémologický, a preto musím začať od úplného základu — od života. Podľa môjho názoru sú naše mozgy osídlené živými organizmami, ktoré jednak v mozgoch ale aj medzi mozgami tvoria živý ekosystém. Ako sa dá tvrdiť o niečom, že je to živé? Ak sa nad tým zamyslíme, tak vlastne nijako definitívne. Nedá sa ako vo fyzike dokázať, že niečo žije. Všetky dôkazy o podstate živého stoja na rôznej miere podobnosti k nám, a my o sebe nepochybujeme, že žijeme. V podstate je teda možný len jeden spôsob dôkazu, a to abduktívny, na základe spoločných podstatných znakov. Známe anglické: ak niečo lieta ako kačica, kváka ako kačica, chodí ako kačica — je to kačica.
Čo sú teda vo všeobecnej (alebo aspoň väčšinovej) zhode znaky
živého? Uvádza sa ich viacero, ale podľa mňa sú najkľúčovejšie
tieto:
- Udržanie stability tvaru pomocou replikácie skôr ako sa tvar rozpadne.
- Udržanie tvaru, zber a modfikácia prostredia pomocou metabolizmu, teda premeny cudzích foriem na vlastné.
- Zámernosť, intencionalita, čiže cielené pôsobenie v smere
predchádzajúcich dvoch znakov života. Vlastne v tej
intencionalite je samotná podstata života, organizovanosť. Ak
rozbijeme bunku na molekuly, z ktorých sa skladá a tie stlačíme
do toho istého objemu, tak žiaden život nebude, iba množstvo
molekúl. Stratila sa intencionalita, organizovanosť. To, že
niektorá molekula práve pozmeňovala tvar ktorejsi inej molekuly,
následne z toho bola reakcia ďalších molekúl atď. To nie sú
vzájomne nezávislé reakcie, ako v neživých systémoch. Každá
reakcia v bunke je v nejakom vzťahu k iným súčasne
prebiehajúcim a je dôsledkom predchádzajúceho toku reakcií.
Teraz si vezmime trebárs príslovie: Komu sa nelení, tomu sa
zelení.
- Nejako sa muselo dostať do väčšiny hláv na Slovensku. Čiže sa
replikuje, čím udržuje svoju stabilitu aj po smrti nositeľa.
- V dotknutých hlavách sa museli nejako premeniť, (metabolizovať)
reakčné dráhy tak, aby keď mi niekto povie Komu sa nelení … v
hlave mi okamžite nastala reakcia: … tomu sa zelení.
- No a teraz si uvedomme, že v tom mozgu sa táto štruktúra musí udržovať, inak by sa zabudla. A to sa v mozgu skutočne deje. Toto príslovie sa aktívne intencionálne usiluje udržať svoju štruktúru. Snaží sa nájsť situácie, kedy sa rozmnoží. Napríklad ja som teraz hľadal vhodný príklad a toto príslovie si z nejakých dôvodov v mojom mozgu vybojovalo svoju priroritu. Z hľadiska toho ako sa to deje v mozgu si prezo muselo príslovie metabolizovať celý rad reakcií od dovtedy nezapojených synaptických štruktúr, pretože príslovie je použité v úplne novej situácii, ako bolo povedzme v detstve keď sa zreplikovalo do mojej hlavy.
Takto som zrazu začal vidieť mémy, keď som sa pustil do štúdia
neurovied. Mozog je vlastne akýsi humus, akási pôda pre život.
Kľúčom k pochopeniu je, že mozog je reaktívny systém. Vlastne je
to strašne husto a neprehľadne zosieťovaný systém nespočetného
množstva reakčných oblúkov: podnet⇒ reakcia.
Metabolizmus
Základom je bunka, neurón. Tá má vstupnú časť, sieť dendritov, čo sú výstupky bunkovej membrány posiate prechodovými kanálmi, a jeden výstupný axón, čo je opäť na konci rozvetvený výbežok membrány bunky. Neurón je čisto pasívny reakčný bod. Funguje takto: Na povrchu membrány udržuje elektrický potenciál -60 mV. Ak cez kanály v dendritoch prúdia kladné ióny, tak sa záporný potenciál znižuje až kým nedosiahne hladinu -20 mV (tzv. akčný potenciál), ktorý spôsobí, že axón začne na svojich výbežkoch produkovať váčky s neurotransmitermi (vazikuly). Medzi dvoma neurónmi je tesný kontakt s úzkou medzibunečnou medzerou, nazývaný synapsia. Synapsia sa skladá z presynaprického neurónu, teda vysielajúceho a postsynaptického, toho, čo prijíma. Ak začne z axónu presynaptického neurónu prúdiť neurotransmiter, ktorý otvorí v postsynaptickom dendrite nejaký prechodový kanál, začne v postsynaptickom neuróne klesať membránový potenciál až na hladinu akčného potenciálu, a postsynaptický neurón začne robiť to, čo predtým predsynaptický neurón jemu: vysielať neurotransmitery zo svojho axónu.
Ak sa zamyslíme, tak vlastne neurón vôbec netuší čo robí a prečo.
Preň ako bunku to nemá nijaký životný význam. Je to preň iba krutá
záťaž, pretože na takúto činnosť musí udržovať a vytvárať bohatý
molekulový aparát. Nie je to súčasť jeho intencionality. Lenže
nedá sa nič robiť, vyššie sme ukázali, že nejako sa práve cez
tieto neuróny musí intencionalita príslovia zabezpečiť. Kde a v
čom? Je ľahké si domyslieť, že intencionalita je v
štruktúre neurónov, v štruktúre synaptických prepojení.
Nechcem zachádzať do podrobností, ono je to v detailoch dosť
zložitejšie, ale v princípe je to o štruktúre nervových prepojení.
Každá informácia si v mozgu musí vedieť zabezpečiť sformovanie
synaptických prepojení, čiže kedy a kde potečie nejaká jej čiastka
ktorými neurónmi. A čo je to informácia? No predsa štruktúra
niečoho, usporiadanie, organizácia. Organizmus je z gréckeho
organon, čo je nástroj, ale aj ústroj, teda niečo nástrojom
vytvorené. V staršej češtine sa organizmus prekladal ako
ústrojenec, niečo ustrojené, stavané, usporiadané.
Kedže je neurón iba reakčný uzol, tak z pohľadu informácie iba
premieňa tok informácie, metaboluje ju. Metabolizmus je z gréckeho
meta (cez) a balló (hádzanie), čiže prehadzovanie niečoho cez
niečo, preskupovanie. V molekulárnom svete sa preskupujú molekuly,
ich spojenia a ich konformácie. V prostredí mozgu sa preskupujú
reakčné toky. Máme tu jeden zo základných rysov živého —
metabolizmus. V mozgu neustále pretekajú toky reakcií aj keď
spíme, či sme v bezvedomí. Len možno spomalene. Mozog nemôže s
týmto prestať, pretože ak by prestal, už nebude vedieť pokračovať,
podobne ako je to v bunke o ktorej som písal vyššie, keď v nej
zastavíme tok procesov. Cez zmysly neustále tečie prúd podnetov.
Tak isto z tela. Tie sú metabolizované ďalej v ďalších štruktúrach
mozgu, a ďalej a ďalej. A zasa v tej istej hierarchii naspäť. V
mozgu neustále prebiehajú vlny reakcií, podobne ako v bunke
neustále prebiehajú prúdy molekulových transformácií.
Napríklad v každej bunke nášho tela (eukaryotickej) v každom momente prebieha naraz 10 tis. exprimácií génov. Exprimácia génu je zložitá reťaz následných krokov, na konci ktorej je konkrétny proteín v konkrétnej konformácii. Ja si ani všetky tie kroky nepamätám. Najprv sa samozrejme musí nájsť príslušný gén na DNA, čo sprostredkuje signálny proteín konkrétneho tvaru. Nájdený gén sa musí zo skrúteného tvaru DNA odbaliť, čo vykoná nejaký iný proteín. Následne sa na začiatok génu napojí iná molekula, ktorá skopíruje gén do prenosovej RNA. Na ňu sa prilepí iný proteín, ktorý zabezpečí rozstrihanie génu na kódujúce a nekódujúce úseky, Kódujúce spojí další proteín do jedného celku, ovšem pokiaľ sa nezapoja iné RNA molekuly, ktoré zabezpečia správne prehodenie kódujúcich častí na konečný výsledný kód. Hovorí sa tomu editácia génu. Ako vidno neustále sa metabolizuje nejaké usporiadanie molekúl. Hotový kód sa dopraví do molekuly ribozómu, ktorý na jeho základe začne syntetizovať z amínokyselín lineárny tvar výsledného proteínu. Tento sa následne veľmi často sám poskladá do funkčného výsledného 3D tvaru, ale ešte častejšie sa počas syntetizovania pripájajú rôzne iné RNA molekuly, či proteíny, ktoré zabezpečia želané skrútenie výsledného tvaru proteínu do funkčnej 3D formy. Preto z jedného kódu môže byť viacero funkčne rôznych výsledných proteínov. Kedy ktorý? Nuž to je veľká otázka, čo a ako rozhoduje. V podstate to vyzerá tak, že rozhoduje celková doterajšia história bunky, ktorej výsledný stav je vlastne intencionalita, organizovanosť v danej chvíli. Máme tu súčasne 10. tisíc lineárnych hierarchických procesov, ktoré sú vzájomne sieťovo previazané plus táto previazanosť je daná historickým nastavením, predchádzajúcim stavom bunky.
Podobné niečo vidíme aj v mozgu. Neskutočne podobné, až mi pri
tom padla sánka. Skúsim to aspoň čiastočne priblížiť. Najlepšie ak
začnem určitým zmyslovým vnemom. a zasa najplepšie bude vizuálnym.
Jednak je pomerne dobre preskúmaný a tiež na ňom som jav najlepšie
pochopil ja.
Vizuálny zmyslový vnem začína na očnej sietnici, tvorenej na svetlo
citlivými bunky. Nad vrstvou týchto buniek je vrstva neurónov, ktoré
sú usporiadané do uzlov, ganglií. Ganglión prijíma signály
z určitého poľa pod ním ležiacich svetlocitlivých buniek, nazývame
to receptívne pole. Každý ganglion z receptívneho poľa vyrobí
výsledný signál. Ale čo vyjadruje tento výsledný signál? Ganglion
spriemeruje intenzitu signálu z každej bunky receptívneho poľa, z
ktorého prijíma signály a tento priemer porovná so susednými
gangliónmi. Von z neho vychádza signál, ktorý reprezentuje
spriemerovaný kontrast receptívneho poľa voči ostatným
susediacim gangliónom. Vec je ešte skomplikovaná tým, že máme dve
sady týchto gangliónov, jedna spracováva menšie receptívne polia a
slúži na tvarovú informáciu o zrakovom vneme, druhá sada zbiera
signály z väčších receptívnych polí a slúži na vyhodnocovanie
pohybu objektov na sietnici. Ale toto nechajme ako zaujímavosť pre
ďalší výklad to nie je podstatné. Rovnako ako to, že takto
vzniknutý výsledný nerv sa všelijako rozdeľuje a kombinuje z
výstupmi z druhého oka.
Tu sa na chvíľu zastavme pri tej podivuhodnej skutočnosti, že očný nerv neprenáša obraz sietnice, ako povedzme CCD chip v kamere nášho mobilu, ale sadu kontrastov receptívnych polí. Ganglión metabolizuje malú časť sietnice na výslednú hodnotu: kontrast voči susedným receptívnym poliam. Vidíme tu základnú jednoduchú transformáciu reakcie očnej sietnice na úplne inú reakciu: kontrast voči iným receptívnym poliam. Táto transformácia, je z pohľadu vstupujúcej informácie, teda rozloženia svetla na sietnici, meta-balló, prehádzaná, preusporiadaná na úplne inú informáciu, metabolizovaná. Takéto elementárne preusporiadanie v jednom uzle metabolických transformácií v mozgu som nazval mém. Mém sa môže a nemusí kryť s nejakou anatomicky definovateľnou grupou neurónov, ako je v oku v podobe gangliónu. Faktom však je, že mozog je celý zložený z neurónových grúp a zrejme väčšina mémov je spojená s týmito elemtárnymi procesnými jednotkami. Už teraz bude užitočné pripomenúť, že aj v bunkách celý rad metabolických zmien prebieha v samostatných metabolických dráhach. Anton Markoš vo svojej skvelej knihe Evoluční tápání pôsobivo argumentuje v prospech samostatných metabolických procesov v zemskej kôre, ktoré mohli byť predchodcom života, keď sa postupne zosieťovali a spojili. Následne sa naučili prekonať smrť tým,. že si do RNA molekúl zapísali svoje tvary a stavy. Nejako takto sa postupne mohol vynoriť bunečný život.
Očný nerv ústi do jedného z mnohých jadierok v thalame,
nazývaného lateral geniculate nucleus (LGN). Pôvodne sa
predpokladalo, že LGN je iba akýmsi retranslátorom do mozgovej
kôry, ale dnes vieme, že 90% vstupov do LGN je z rôznych oblastí
mozgovej kôry, teda, že sa aj v LGN sa nachádza množstvo mémov,
ktoré zrejme robia nejaké metabolizácie spojené s vyššou mozgovou
činnosťou. Inak povedané: to čo vidíme sa modifikuje inými
oblasťami mozgu.
Z LGN potom ide signál do mozgovej kôry, do oblasti nazývanej V1.
Tá je opäť zložená z obrovského množstva grúp neurónov. Úlohou
tejto oblasti je transformovať vstupujúce kontrasty receptívnych
polí do hrán, čiar, ktoré reprezentujú tvarovosť očného vnemu.
Napríklad tvár má obrys, vnútri linky úst, očí nosa atď. Ako je
oblasť V1 organizovaná? Vezmime si napríklad niekoľko susediacich
receptívnych polí, reprezentovaných kontrastami medzi nimi.
Signály týchto kontrastov súčasne vstupujú do veľa susediacich
grúp vo V1. Každá z týchto grúp je vnútri seba náhodne rôzne
poprepájaná tak, že reprezentuje určitú orientovanú čiarku. Teda
ne jednu skupinu vstupných kontrastov je veľa možností ako je tá
čiarka orientovaná. Povedzme po pár stupňoch okolo osi skupiny
receptívnych polí. Tá skupina neurónov, ktorej štruktúra najlepšie
zodpovedá tvaru čiary zloženej z kontrastov receptívnych polí
vydáva najintenzívnejší výstupný signál do ďalších oblastí mozgu.
Povšimnime si niekoľko detailov tohoto javu. V prvom rade je
vidno, že informácia opäť metabolizuje do nejakej inej. Z
kontrastov na elementy tvaru obrazu. Ďalej, že je to robené
mémami, teda elementami metabolizmu. No a napokon, že súčasne
pôsobí veľa mémov na urobenie toho istého. Samotný jeden mém nevie
čo robí, a nevie či vyhodnotil správne vstupujúce kontrasty, iba
to posiela ďalej. Jeho reakcia je povedzme v škále od 1-10 na
úrovni 5, susedného mému 8 a niektorého dokonca 9. To, či jeho
vyhodnotenie sady kontrastov najviac zodpovedá nejakému tvaru sa
rozhodne až niekde ďalej v inej oblasti mozgu. Podobne aj v
bunkách prebieha veľa metabolizujúcich procesov, ktoré nemusia
nevyhnutne zodpovedať momentálnym potrebám bunky a ich metabolity
sa následne likvidujú.
Z V1 vystupujú signály do oblasti V4, kde sa vyhodnocuje pohyb na sietnici a do V2, kde sa zapamätávajú tvary. V podstate je V2 súčasťou javu, ktorý je v kognitívnych a neurovedách nazývaný faktickou pamäťou. V oblasti V2 sa zapamätajú tvary niečoho zložené z čiar v oblasti V1. Teda jedna grupa vo V2 mapuje veľa grúp vo V1 a grupy vo V1 ústia do veľa grúp vo V2. Napríklad tvar tváre matky reprezentuje nejaká grupa vo V2. Oblasť V2 sa formuje učením. Z pohľadu mojej interpretácie tu hovoríme o mémoch, ktoré sa zreplikovali z prostredia. Teda elementárne reakčné útvary, elementárne metabolické dráhy zložené z následnosti metabolických transformácií informácie vstupujúcej do očí. Štruktúra svetelných lúčov vstupujúca do oči sa postupne metabolizuje na tvar zložený z obrysov.
Čo rozhoduje o tom, či práve toto mapovanie grúp z V1 je to
správne a nie iné? Ak si to predstavíme, tak súčasne vo V2 v
rôznej miere veľa grúp reaguje na rovnakú sadu reakcií z V1.
Rozhoduje odpoveď z prostredia. Mozog nielen prijíma informácie
ale v rovnakej miere reaguje a pôsobí na prostredie. Dieťa po
narodení neustále skúša všetky možnosti. Chyba značí nesprávnu
metabolická dráha, úspech značí správnu metabolická dráha.
Napríklad fixácia na tvár matky je súčasne riadená opticky ale aj
olfaktoricky pachom. Dieťa má zapamätanú vôňu mlieka od matky
spojenú s jej tvárou. Tvar tváre matky sa postupne dopĺňa o ďalšie
a ďalšie detaily. napríklad z rôznych uhlov. Takto sa postupne v
oblasti V2 niektoré synaptické spojenia posilňujú iné rušia.
Navyše dnes dokázatelne vieme , že akýsi základný tvar tváre je
vrodený, čo isteže uľahčuje prvotný výber medzi grupami vo V2.
Ďalej sa ukazuje, že všetky podobné tvary sa vo V2 nachádzajú
anatomicky vedľa seba. Mémy tvarov sa vyskytujú v
širokej variabilite, tak aby boli schopné zmetabolizovať prakticky
všetky možné tvary do nejakého nejbližšieho vnútorne
metabolizovaného tvaru. Inak povedané, tvary, ktoré vidíme sa v
skutočnosti môžu líšiť od toho, ako sú v mozgu metabolizované, ale
skúsenosť reprezentovaná v iných komplexnejších mapovaniach mozgu
to skoriguje na tú najpravdepodobnejšiu vnútornú metabolickú
dráhu. Pripomeňme si, že 90% vstupov do LGN ide z rôznych oblastí
kortexu, ktoré takto môžu modifikovať zrakový vnem podľa skúsenosti. V
mozgu sa reakcia vždy konštruuje, nezapamätáva sa.
Všimnime si, čo rozhoduje o tom, ktoré mapovania sú správne a ktoré nie. Prírodný výber. Tá štruktúra metabolitov z V1, ktorá najviac zodpovedá skúsenosti z prostredia vyhrá v konečnom mapovaní vo V2. Dá sa predstaviť, že tak ako vo V1 aj vo V2 sa v procese učenia môžu vyskytnúť mémy, ktoré nikam nemetabolizujú. Mozog totiž pri svojom prvotnom formovaní v maternici naseká v každej svojej oblasti nadbytok grúp neurónov. Také, ktoré sú mozgu nanič sú potom likvidované, samy apoptujú. Mozog novorodenca má myslím o 20% viac neurónov ako ho má v puberte. Všimnime si to najpodstatnejšie: mozog obsahuje intencionalitu svojho nositeľa iba v základnej svojej štruktúre, ale intencionalitu svojho obsahu neobsahuje. Informácie, ktoré učením formujú mozog majú intencionalitu vlastnú, inak by predsa neboli schopné vytvoriť zodpovedajúce štruktúry reakcií. Napríklad tvar krychle je od vrodenej intecionality organizmu nezávislý, na jeho rozpoznanie sa musí v mozgu vytvoriť zodpovedajúca metabolická línia. Mozog nemá nič vrodené k tomu aby rozhodol že toto je krychla a toto válec. Zato tvar krychle má svoju tvarovú intencionalitu, schopnú tak sformovať vrodené prostredie mozgu k reakcii toto je krychla, alebo toto válec. Informácia tvaruje mozog, nie naopak, že by mozog tvaroval informáciu. Je vcelku podivuhodné, že tento jednuchý postreh systematicky chýba aj neurovedcom.
Poďme však ďalej. Vo vizuálnej časti kortexu sú ďalšie oblasti, ktoré následne vo svojich grupách-mémoch metabolizujú tok reakcií z oblastí V2 a V4. sú tam oblasti, ktoré doplnia plochy medzi čiarami o farbu a celý rad ďalších. Dôležité je všimnúť si, že tu prebieha hierarchická následnosť metabolických premien. Kde pokračuje táto hierarchia?
Podobné hierarchické metabolické toky sú zo všetkých zmyslov:
somatozenzorických, olfaktorických a sluchu. Je tu však aj výstup
z mozgu, ktorý spravidla ústí do ovládania skupín svalov, teda
motorická hierarchia, ktorá je opačne otočená. Najbstraktnejšia
úroveň je v najvyššom bode hierarchie, ale z hľadiska
metabolizačného toku je na začiatku. Tam sú naučené koordinácie
svalov na nejaký konkrétny pohybový ciel, napríklad pohnúť rukou
hore. Pod tým sú tie konkrétnejšie, teda ktoré svalové partie sa
zapoja, no a to ústí cez miechu do ganglií pri jednotlivých
svaloch, ktoré robie podobné niečo ako ganglie v oku, dráždia
nejaké svalové pole. K tomu treba prirátať malý mozog, ktorý
toto všetko dolaďuje na jemnú motoriku.
Z doteraz povedaného môžeme vyvodiť kľúčový poznatok: toky informácií zo zmyslov sú metabolizované v slede následných mémov usporiadaných do hierarchie. Podobne zostupuje motorická reakcia organizmu v hierarchii metabolizovaním hlavných motorických koordinácií na stále podrobnejšie koordinácie svalov až do svalov samotných. V terminológii neorovied tu máme teda vzostupné dráhy spracovania senzorických informácií a zostupné dráhy spracovania motorických informácií.
Obe tieto triedy metabolických transformácií sa spájajú v tzv.
asociatívnych oblastiach kortexu. Zdá sa, že primárnym významom týchto oblastí je dotvorenie
konečného reflexného oblúka: podnet z prostredia ⇒ svalová reakcia
na prostredie. Aj asociačné oblasti sú tvorené grupami neurónov,
ktoré sú vnútorne všelijako prepojené (mémy) a husto prepojené navzájom.
Vstupujú sem metabolické výsledky zo spracovania zmyslových
informácií a vystupujú odtiaľto prepojenia na primárne motorické
oblasti. V asociačných oblastiach sa môžu vzájomne prepojiť
zmyslové vnemy do jednej udalosti spojenej so správnou motorickou
reakciou na túto udalosť. Napríklad pohyb na okraji zorného poľa
spojený s charakteristickým zvukom spôsobí otočenie hlavy za
zdrojom zvuku a obrazu.
Do tejto ešte pomerne prehľadnej a zrozumiteľnej schémy však
zásadne zasahujú ďalšie časti mozgu, a to podkôrové oblasti a
prefrontálny kortex. Významné pre nás sú najmä bazálne gangliá,
hippocampus, thalamus a amygdala. Hippocampus je akoby centrom metabolických tokov, pretože cezeň sa všetko prepája so všetkýc v bohatých
vzostupných a zostupných dráhach.
Hippocampus je zodpovedný za vytvorenie dlhodobej pamäte. Všetko čo, sa nám aktuálne deje ide zo zmyslových oblastí aj do hyppocampu a odtiaľ všade do mozgovej kôry a aj do podkôrových oblastí a starších oblastí (stredný mozog). Nový tok do hippocampu si hľadá najpravdepodobnejšie prietoky do všetkých kórových oblastí, a keď už existujú nejaké metabolické dráhy, ktoré sa najviac podobajú práve prechádzajúcim tokom zo senzorov a z ich asociácií v asociačbnej oblasti, tak to hippocampus pozná zo spätných reakcií a od toho momentu udržuje tento reflexný oblúk dráždenia v neustávajúcom krútení. To je dobré k tomu, aby si nové toky cez synapsie stihli zúčastnené neuróny upevniť, čo nejakú dobu trvá, pretože na to potrebujú posilniť v patričných dendritoch nové kanály a zodpovedajúci molekulárny aparát. Ak nový podnet nie je významný na základe doterajšej skúsenosti, tak takto vytvorený excitačný oblúk hyppocampus⇄kôra je inými oblasťami mozgu tlmený, čoho výsledkom je, že sa nezapamätá.
Správna otázka je, no ale ako vie mozog, čo je významné a čo nie? Teda, čo rozhoduje o tom, ktorá udalosť sa má zapamätať a ktorá nie? Odpoveďou je podľa mňa vrodený hodnotový systém organizmu. Nesmieme zabúdať, že jediný zmysel života každého organizmu v darwinistickom svete je žiť pre vlastné rozmnoženie dovtedy, kým sa nerozpadnem. K tomu musím zbierať zdroje, chrániť svoju integritu a u pohlavne sa množiacich druhov hľadať partnera na párenie, prípadne investovať zdroje do potomkov. Tieto základné úlohy musia byť dáko v mozgu zakódované vo vrodených mémoch, teda vrodených metabolických uzloch.
Pokiaľ je mne známe, tak všetky hodnotové metabolizovania sú v modifikovné v amygdale. Amygdala je cez hyppocampus, bazálne gangliá, thalamus a hypotalamus bohato prepojená, a tak isto je prepojená s prefrontálnym kortexom. V amygdale sa do nej prúdiacim metabolitom prisudzuje jedna z rekacií na ohrozenie: uteč, útoč, stuhni, podvol sa. Myslím, že vyhodnotenie rizík podnetov a prisúdenie im patričnej základnej orientácie výsledných metabolických tokov reakcií je riešené grupami neurónov v amygdale. K tomu sa pripájajú aj spomínané ďalšie podkôrové oblasti, teda bazálne gangliá a thalamus. Napríklad niektoré jadierka v bazálnych gangliách riadia pohyb očí podľa situácie, pretože v spolupráci s amygalou vytvárajú pozornosť, koncentrovanie sa na výber dôležitých metabolických tokov.
Na záver tejto prehliadky mozgu uvediem prefrontálny kortex, ktorý je opäť bohato prepojený hádam so všetkými oblasťami v mozgu. Je tvorený mnohými jadierkami špecializovanými na nejaké typy úloh a v tých jadierkach opäť nájdeme grupy neurónov, ktoré, sú vnútorne zrejme náhodne poprepájané, ako tvrdí autor neurálneho darwinizmu Greg Edelman. Tvrdí sa, že úlohou PFC je koordinovať všetko v mozgu, vytvárať modely situácií zložené z doterajších skúseností, a na základe týchto variantných modelov rozhodovať čo najbližšie vykonám. Čiže plánovanie. Ak sa na vec pozrieme z pohľadu mémového, tak PFC je okoby taký integrujúci metabolizér všetkých výsledkov metabolizmu. V ňom sa stretávajú a vzájomne potláčajú všetky možné súčasne pôsobiace metabolické toky, pretláčajú sa kompetujú, symbiujú a výsledok posúvajú ako modifikáciu celkového správania. Zrejme to, čo nazývame vedomím, má podstatnú informačnú funkcionalitu spojenú s PFC.
Potiaľ veľmi zjednodušený a stručný model toho, čo asi v mozgu
prebieha. Pozrime sa na to teraz z pohľadu živosti.
Z vyššie napísaného myslím jednoznačne vyplýva, že z pohľadu
pôsobenia prostredia vstupujúceho do mozgu a z pôsobenia na
prostredie vytupujúceho z mozgu v podobe svalových činností,
hovoríme o neustálej transformácii neurónových reakcií. Objekty
prostredia vstupujú do mozgu ako tvary, znaky, teda informácie o
prostredí a okamžite za hranicou tela sa transformujú na reakcie.
Informácia sa transformuje grupami neurónov na úplne inú podobu.
Jednotlivé kroky tejto transformácie, ktoré som nazval mémami, z
pohľadu podstaty veci abduktívne dokazujú metaboličnosť celého
procesu. Máme tu molekuly (štruktúry synapií) a máme tu
metabolické kroky transformácie z jednej podoby molekúl na inú
(reakcia grúp neurónov na vstupné toky a následné dráždenie
ďalších grúp). Máme tu transformačné línnie ako sériu
metabolických krokov, metabolické dráhy mémov. A máme tu dokonca
metabolické slučky, okruhy premien metabolitov, ktoré sa neustále
vracajú na svoj začiatok. Jeden taký druh metabolický kruh sme
popísali ako proces premeny dočasnej pamäte na trvalú.
Metabolických kruhov je v mozgu veľké množstvo. Vďaka novým zobrazovacím metódam sa podarilo zistiť veľmi dôležitú vec. Nielenže sú metabolické dráhy zostavené priamým prepojením axónami medzi oblasťami, ale pri tak hustom prepojení mozgových oblastí sa vytvárajú metabolické dráhy na konkrétne typy reakcií. Boli objavené tzv. Large Scale Networks. Doteraz ich bolo objavených myslím do stovky. Najznámejšie sú tri: defaultná, salientná a exekutívna sieť. Exekutívna sieť sa vytvára, keď mozog interaguje s prostredím, defaultná vtedy, keď v sebe myslíme, ale nekonáme von, a salientná rozhoduje, kedy je aktívna exekutívna a kedy defaultná. Práve v poruche salientnej siete hľadajú príčiny paranoí a schizofrénií, kde mozgy nie sú schopné odlíšiť to, čo je v hlave a mimo hlavu. Z nášho pohľadu však je dôležité si uvedomiť, že mémy nemusia byť lokalizované na grupy neurónov. Mém vždy vznikne, keď sa vytvorí nejaký nový metabolit, teda nejaké usporiadanie synaptických prepojení v mozgu. Mém je kúštik informácie, nie kúštik mozgu.
Dúfam, že som predložil dosť argumentov pre jeden aspekt života v mozgu, pre metabolizmus. Teraz si skúsme predstaviť mozog ako svet, kde neustále tečú metabolické toky, vzájomne sa prelínajú, menia, zlučujú, štiepia atď. Z metabolických dráh sa vytvárajú metabolické cykly, metabolické siete a hierarchie. Spomínam opäť knihu A. Markoša Evoluční tápaní. Niečo podobné podľa neho pred 4 mld rokov prebiehalo v zemskom plášti. Taktiež tam boli silikátové štruktúry, ktoré „vrodene“ usporiadavali tieto toky podobne, ako máme v mozgoch vrodené prepojenia mozgových oblastí a vlastne celú anatomickú architektúru v mozgu. Zároveň sa nedá odolať pripodobneniu tohoto diania k dianiu v bunkách. Život v bunke je podobne neustály organizovaný prúd metabolických dráh, sietí a cyklov, ktoré sa vzájomne ovplyvňujú. Dnes sa ukazuje, že aj baktérie budujú veľmi zložitý systém vzájomných vzťahov založený na vzájomnom preberaní a metabolizovaní látok a na horizontálnom toku génov. Ktoho vie, zdá sa, že živé systémy sú primárne o metabolických sieťach svojich nositeľov a až sekundárne o rozmnožovaní.
Intencionalita
Zamyslime sa teraz nad intencionalitou (zámernosťou) živého. Ako
sa vlastne vybudovala, ako funguje? Organizmy (teda tie
proteínové) aktívne vyhľadávajú potrebné zdroje, cielene ich
transformujú na komponenty potrebné pre vlastnú štruktúru,
vylučujú tie nepotrebné či škodlivé, u pohlavne sa množiacich
aktívne hľadajú partnerov a tiež sa cielene starajú o potomkov.
Nezávisia (ako napríklad kryštalické štruktúry) od priazne
prostredia a svoju replikáciu si na prostredí vymáhajú. To všetko
sa muselo vyvinúť z jednoduchých začiatkov. Podľa mňa je takým
jednoduchým príkladom intencionality metabolický cyklus, napríklad
Krebsov (citrátový) cyklus. Je to v kruhu s opakujúca premena
molekúl, pri ktorej sa cielene vypúšťajú a pripájajú molekuly z
prostredia. Cyklus sám seba udržuje, kým má dostatočný prísun
vstupujúcich molekúl v prostredí. Niečo iné je prostá séria
metabolických reakcií, ktorá iba začne a skončí na svojom konci.
Metabolický cyklus nemá vlastne koniec a v jeho cykličnosti je
jeho intencionalita.
Už v tomto bode si nemožno nevšimnúť paralely v mozgu. Vyššie som
spomenul cyklus medzi hippocampom a oblasťami v kortexe, pomocou
ktorého sa z krátkodobej pamäte vytvára dlhodobá. Takýchto cyklov
je v mozgu obrovské množstvo. Vstupujú do nich podnety z iných
oblastí mozgu a vystupujú podnety do ďalších oblastí. Niekde
reakcie vytvárajú inde tlmia. Vlastne sa dá povedať, že dlhodobá
pamäť je v trvalom nastavení tohoto cyklu. Podobné cykly sú cez
kortiko-bazálne gangliá-thalamo-kortikálne slučky (Cortico-basal
ganglia-thalamo-cortical loop), v ktorých sa zrejme udržuje emočné
nastavenie voči podnetom. Ak je v niektorom uzle tejto slučky
deviácia, tak sa objavuje celá škála porúch nálad, napríklad
hystérie či depresie, afektívne poruchy.
Ďalej sa (myslím) dá vidieť intencionalita vo vzájomnej
previazanosti jednotlivých metabolických dráh a cyklov. Napokon
metabolické zmeny sú závislé od vstupov, ktoré odniekiaľ musia
vstupovať (z metabolických dráh či cyklov) a majú výstupy, ktoré v
živej bunke niekam konkrétne mieria, slúžia na niečo, vstupujú do
iných metabolických dráh alebo cyklov.. Napokon v čom inom, ak nie
v tomto, je živosť, organizovanosť, samo-usporiadanosť?
V mozgu vidíme to isté. Veľká previazanosť jednotlivých oblastí
mozgu a vzájomná previazanosť grúp neurónov v oblastiach
spôsobuje, že každá grupa neurónov, ba každý neurón môže vstupovať
do mnohých metabolických dráh reakcií. Doslova môže byť súčasťou
viacerých mémov (vo vyššie uvedenej definícii) súčasne. Takú
mimoriadnu vnútornú organizovanosť nedosahujú ani eukaryotické
bunky. Veľká previazanosť v mozgu je predmetom pokračujúceho
výskumu a zďaleka mu niet konca. V prípade hlbšieho záujmu treba
googliť brain connectome.
Ďalší prejav intencionality je aktívna znalosť prostredia a aktívne pôsobenie na prostredie. Množstvo metabolických procesov v bunke začína vstupom molekuly z prostredia, buď ako vstupná látka do metanolických línií, alebo ako signálna látka, ktorá spustí reťaz vopred pripravených metabolickýcj línií. Napríklad vyšše som uvádzal, čo sa deje zhruba pri exprimácii génu, tak tento proces môže začať vstupom signálneho proteínu na membránu bunky, ktorý jej oznamuje, že má vyrobiť nejaký výsledný proteín. Intencionalita je v selektívnosti procesu. Bunka reaguje iba na tie podnety z prostredia, na ktoré má vnútorné metabolizačné schémy a pôsobí voči prostrediu z rovnakých funkčných príčin.
V podstate veľmi podobné procesy vidíme aj v mozgu. V popise spracovania vizuálneho vnemu som sa vyššie snažil popísať ako sa od prvej transformácie fyzikálneho vnemu na metabolit kontrastu receptívneho poľa spúšťa metabolický rôzne sa vetviaci a sieťovaný prúd reakcií a transformácií do iných metabolitov,, ktoré tvoria vnútornú intencionalitu sveta neurónových synapsií. Je v tom selektívnosť, pretože (ako sme videli) tvar vstupujúci do očí nemusí zodpovedať výslednému tvaru, ktorý sa vyselektuje vo vnútornej štruktúre. Že je tomu tak dosvedčuje celá plejáda očných klamov.
Tretí rys intencionality živého je spojený so snahou udržať stabilitu svojho tvaru replikáciou do pokiaľ možno úplne rovnakej štruktúry. Kedže tento rys súvisí s rozmnožovaním živých systémov, popíšem to v nasledujúcej kapitolke eseje.
Replikácia
Myslím, že by bolo nosením dreva do lesa vysvetľovať bližšie
význam replikácie pre živé systémy. Treba snáď len pripomenúť, že
tento rys živého je nositeľom darwinovskej evolúcie. Replikácia s
chybami, s variantami vedie v prostredí s obmedzenými zdrojmi k
prírodnému výberu foriem s najúčelnejšími variantmi znakov.
Interakcia organizovanej metabolickosti s prostredím je formujúcim
činiteľom všetkého v darwinovskom svete. Organizmy nielen, že sú samy o sebe,
ale sú vždy a zakaždým neoddeliteľnou súčasťou prostredia, ktoré
ich formuje, a ktoré spoluformujú ony. Darwinizmus bez ekológie
nedáva zmysel.
V úvode som ako príklad replikujúceho sa systému uviedol
príslovie. To však je už dosť zložitá vec na vysvetlenie toho, ako
asi príslovie žije v mozgu. Svoj pohľad na vec sa pokúsim
vysvetliť na jednom slove.
Jazyk je pochopiteľne odvodený od sluchového systému. Ten začína
v uchu, kde sa zvuk rozloží na frekvenčné podiely. Pokračuje v
jadierkach thalamu, kde metabolizuje na rôzne reakčné entity,
napríklad na mémy časového rozdielu dopadu signálu do oboch uší,
mémy frekvenčných rozdielov a celý rad ďalších. Metabolické dráhy
potom pokračujú do primárnej zvukovej oblasti kortexu zvanej A1.
Tam údajne zatiaľ dosť tápeme, prebieha tam paralelne mnoho
metabolických línií. Jazyk sa z A1 vytvára v dvoch slávnych
oblastiach v ľavej hemisfére: v Broccovej oblasti a vo Wernickovej
oblasti. V Broccovej oblasti sa pravdepodobne rôzne aspekty zvuku
metabolizujú do základných elementov jazyka, do foném, z toho
morfém atď. Vo Wernicovej oblasti sa pravdepodobne z častí
skladajú slová, vety, skladá sa význam transferovaný do iných
oblastí kôry.
Nech už je to ako chce, skúsme si podľa tohoto modelu predstaviť
slovo ako živý systém. Z pohľadu mozgu slovo vstupuje ako v čase
rozložený zvuk, čo je aj takto metabolizované v podobe mémov. Máme
napríklad mémy fonémov, čo sú základné zvukové elementy jazyka.
Slovo je zložené z mémov-fonémov. Aby tomu tak mohlo byť, počas
prvých mesiacov života sa v mozgu v Broccovej oblasti vytvárajú v
grupách neurónov primárnej oblasti A1 všelijaké patterny zvukov
podobne ako vo vizuálnej V1 patterny orientácie čiar. Z týchto
vzorcov zvuku sa v Broccovej oblasti zachycujú najčastejšie
jazykové elementy - fonémy. Dieťa žije obklopené určitým jazykovým
prostredím a fonémy materinského jazyka ho najčastejšie obklopujú.
Ako Broccova oblasť vie, že ide o jazyk a nie iný typ zvukov
neviem, zrejme pôjde o prepojenie týchto oblastí cez asociačné a iné v mozgu. Napokon Noam Chomski si urobil slávu na tom, keď dokázal, že
musíme mať nejaké vrodené jazykové štruktúry v mozgu, pretože
všetky jazyky majú spoločnú základnú štruktúru.
Pozrime sa na tento proces z pohľadu mémov. Do uší dieťaťa
vstupuje zvuk v určitej štruktúre, ktorá je daná jazykom jeho
prostredia. Tento tok si cez vrodené metabolické štruktúry mémov
jazyka formuje na grupách Broccovej oblasti mémy, ktoré najlepšie
odrážajú metabolické transformácie mémov z ucha cez vrodené mémy
jazyka. Prvé, čo si musíme na celom procese všimnúť je jeho
nesporná intencionalita. Slovo si vytvára a modifikuje reakčné
toky v mozgu tak, aby zodpovedali jeho štruktúre. To, čo malo
pôvodne fyzikálnu formu chvenia vzduchu na ušnom bubienku v mozgu
nadobúda intencionálne samoudržovanú štruktúru mémov vo vzájomných
metabolických vzťahoch. Fyzikálna štruktúra chvenia zvuku,
teda znak prostredia bez vlastnej intencionality (nevie sa
sám seba udržiavať), sa mení na význam sám o sebe, teda
štruktúru mémov s vlastnou intencionalitou.
Pokračujme však ďalej. Koncom prvého roku života dieťa začína vokalizovať slová. Pokúša sa z odpočutých fonémov vytvárať asociácie na motorické oblasti hlasiviek, regulácie dýchania a tvarovania úst a hrdla. Pokúša sa slová replikovať. Kedže ide zasa len o formovanie mémov metabolizujúcich reakčné toky z Broccovej a Warnicovej oblasti do asociatívnych oblastí a odtiaľ do motorických mémových oblastí, opäť môžeme hovoriť o intencionalite slova, ktoré sa neustálym upresňovaním metabolických tokov pokúša zreplikovať samé seba. Podstatné je, že slovo nie je ten fyzikálny tvar idúci z úst do ucha, ale štruktúra mémov a ich vzájomný metabolizmus v mozgoch hovoriaceho a počúvajúceho. Nie je pri tom dôležité akú konkrétnu synaptickú štruktúru má vysielajúci mozog a akú má prijímajúci. Dôležité je, slovo si dokáže zabezpečiť, aby v oboch mozgoch prebehla rovnaká metabolická transformácia od ucha dnu a zdnuka von tak, aby vždy zaznela rovnaká fyzikálna akustická štruktúra.
Všimnime si podstatné znaky javu:
- Slovo sa dokáže rozmnožovať.
- Slovo existuje v mozgu v podobe organizovanej štruktúry metabolických transformácií mémov.
- Slovo sa rozmnožuje formou artefaktu prostredia, teda
fyzikálne ako zvuk.
Z tretieho bodu vyplýva, že artefakt prostredia slúži ako dočasný zápis nejakej štruktúry synapsií mimo mozog. Nedá sa nevidieť paralelu s proteínovým životom, kde podobnú úlohu plní RNA a DNA, do ktorých je štruktúra proteínov zapísaná a čítaním ktorých sa znova vytvorí nový proteín.
Súčasne je zrejmé, že cez replikáciu vonkajších artefaktov a ich
správnu vnútornú implementáciu sa realizuje Darwinov prírodný
výber. Ten vonkajší artefakt, ktorý sa dokáže správne
implementovať do čo najviac vnútorných synaptických štruktúr je
prírodným výberom uprednostnený.
Domnievam sa preto, že nič v skutočnosti nebráni chápať
synaptické štruktúry v mozgu, ktoré metabolizujú nejaké štruktúry
na iné tak, aby udržali svoju štruktúru a ktoré dokážu túto svoju
štruktúru rozmnožiť, čím si udržujú vlastnú intencionalitu, za živé
a dal som im názov mémbionty. Každé slovo je jednoznačne
mémbiont. Slová sú zrejme súčasne možno najstaršia forma mémbiontov a
tiež najjednoduchšia. Niečo ako najjednoduchšie baktérie.
Iný život
Podobnosť mémového života s proteínovým je podľa mňa až zarážajúca. Súčasne však si treba neustále dávať pozor na to, že ide o úplne, ale úplne inú formu života, a teda aj iné zákonitosti živého.
Prvý dôležitý rozdiel je v tom, že mémový život môžeme považovať za nefyzikálny, nemateriálny. Jeho podstata je čisto informačná, štrukturálna. Iste, vždy ide o štruktúru synapsií neurónov a tie sú fyzické. Lenže ich fyzickosť je súčasťou inej intencionality, tej proteínovej. Slovu je úplne jedno, ktoré konkrétne synaptické štruktúry ho vytvoria a čo to stojí námahy zúčastnených neurónov. Synapsa ako synapsa, je na neuróne aby ju vytvoril podľa štruktúry mémbionta.
Jeden z dôsledkov nematariálnosti mémového života je, že jeho metabolizmus je totožný s jeho intencionalitou a organizovanosťou. V proteínovom živote je metabolizmus materiálny, organizovanosť je organizovanosťou molekúl a závisí od fyzických vlastností molekúl. Aj proteínový život si od tejto závislosti výrazne pomohol svojou stavebnicovou štruktúrou, kde sa z pomerne málo legových kociek, 21 amínokyselín, stavia zhruba 200 miliónov známych proteínov, a tie navyše sú variantné vo viacerých 3D konformáciách s odlišnou funkcionalitou. Predsa len však je proteínový život stále ešte molekulový. Napokon molekulárnosť proteínového života je asi jednou z hlavných príčin pre vytvorenie ochranných obalov organizmov — bunkových membrán, telesných povrchov, slizníc, s následnou nutnou oddelenosťou vnútorného a vonkajšieho prostredia, teda klauzurou.
V mémovom živote je mém štruktúra, ktorá je metabolizovaná štruktúrou iného mému, pričom nevyhnutne nemusí dôjsť ku zmene synaptických štruktúr neurónov. To je vidno v tzv. krátkodobej pamäti (working memory), kde dočasne hippocampus udržuje nejakú štruktúru v mozgu, a keď sa táto následne neutvrdí opakovaním, alebo nejakými ďalšími kontextovými zaradeniami, tak sa zabudne. Takisto sa v mozgu vytvárajú veľkoškálové mémy na riešenie nejakej okamžitej reakcie, pričom sa nezmení synaptická štruktúra. Informácia mení sama seba, pričom nemusí meniť svojho nositeľa.
Jeden z dôsledkov je, že v mozgu je možný skoro akýkoľvek
nemateriálny svet. Fyzikálne obmedzenia sú len v krajných
hodnotách, napríklad mozog nemôže vydať viac energie na zmeny
synapsií, či na nervové prenosy, ako je fyzikálne možné. To je
treba mať na mysli v porovnaní so svetom proteínového života,
ktorý neustále naráža na fyzikálne obmedzenia pri svojom
budovaní.
Významný rozdiel mémového života oproti proteínovému vyplýva z mimoriadnej prepojenosti oblastí v mozgu, teda v hustom mozgovom konektóme. V zásade nič nebráni tomu, aby bola ktorákoľvek časť mozgu nepriamo prepojená s ktoroukoľvek inou. Grupy neurónov ale aj jednotlivé neuróny sa takto neraz súčasne zúčastňujú niekoľkých metabolických procesov naraz. Neviem, či sa dá nájsť niečo podobné v proteínovom svete, kde jednotlivé bunky sú oddelené membránou a telá povrchovými bariérami. Povedzme jeden proteín nemôže súčasne plniť rôzne úlohy v dvoch bunkách. Vo svete mémového života je to pravidlom.
Hlavným dôsledkom tejto skutočnosti je, že kým v proteínovom svete je vytváranie symbióz pomerne zložitá záležitosť, vo svete mémov niet nič ľahšie. Napríklad vyššie uvedené príslovie ako mémbiont endosymbioticky v sebe obsahuje mémbionty slov, z ktorých je zložené. Súčasne tieto slová sú endosymbiontami mnohých iných viet, v ktorých sa vyskytnú buď dočasne, alebo ako súčasť nejakej podobnej zapamätanej sentencie. Slovo komu je súčasťou príslovia Komu sa nelení tomu sa zelení ale aj Komu pánboh tomu všetci svätí. Vďaka tejto mimoriadnej miere schopnosti symbiotizovať sa je vôbec možné mať v hlavách komplexné modely prostredia.
Druhou stranou mince však je menšia schopnosť vytvárať uzavretosť
organizmu, jeho ohraničenosť od okolia, teda to, čo A. Markoš
nazýva klauzura organizmu. Je veľmi ťažké v hustom konektóme
udržať tvar. Napriek tomu to očividne funguje. Naučený tvar
predátora ostane nezmenený prakticky celý život. Je vecou hlbšieho
výskumu zistiť ako sa udržuje hranica medzi stálosťou a zmenou
tvaru mémbiontov. Istotne jedným z hlavných faktorov, ktoré na to
pôsobia v mozgoch ľudí je stálosť artefaktov, ktoré sú génmi sveta
mémov. Hoci náš príklad slova alebo príslovia môže mať v každom
mozgu inú podobu, pri jeho replikácii sa musí zachovať jeho
ustálená podstata v štruktúre artefaktu.
Darwinistický svet mémbiontov
Svet mémov bol darwinistický dávno predtým, ako sa v ňom objavil
replikujúci sa život. Ako sme vyššie ukázali, mozog je reaktívny
systém, svoje reakcie nastavuje a získava v interakcii s
prostredím, v ktorom žije. Rôzne varianty reakcií sú vystavené
selekčnému tlaku prostredia a vyhrávajú tie, ktoré sú pre
organizmus najvýhodnejšie. Veľmi dobre a výstižne to popisuje Greg
Edelman vo svojom koncepte neurálneho darwinizmu. V mémologickej
terminológii nie každý mém uspeje a nie každá metabolická zmena má
rovnakú šancu na prežitie.
Dôležité, myslím, na tom je, že od narodenia je v mozgu selekčné
prostredie, kde súťažia rôzne metabolické dráhy, či rôzne
jednotlivé mémy. Uspejú také, ktoré z prostredia prišli skôr a
našli vhodný kontext, do ktorého sa mohli zaradiť. Tým však
súčasne vytvoria nový kontext, s ktorým musia zápasiť neskôr vstupujúce mémy. Od počiatku je v mozgu preto ekosystém,
nie jednotlivé vzniklé darwinistické entity. Zatiaľ som naklonený
nazývať tento ekosystém egosystémom, hoci mi pri tom
prekáža antropocentrický charakter pojmu ego. Zaujímavé je, že
podľa Markoša život nezačal na Zemi z nejakého posledného
prapredka — LUCA, last common universal ancestor — ale z
ekosystému rôznych spoločných predkov — LECA, last enviroment
of common ancestors. Mozog organizmov mi pripomína z tohoto uhlu pohľadu
štruktúrovaný priestor, v ktorom znova a znova povstáva život po
každom narodení. Znova a znova sa pokúša o evolúciu, lenže tá je u
drvivej väčšiny mozgov ukončená predčasne smrťou hostiteľa.
Mémbionty, ktoré sa v ich mozgoch vyvinuli nemajú podstatnú
schopnosť živého: rozmnožiť sa do iných mozgov.
Aj u iných druhov, ako sme my, však v obmedznej miere vidíme túto schopnosť. Napríklad vtáky sú pri ich nevyhnutne malých mozgoch schopné neuveriteľného výkonu (viď napríklad vrany v jednom japonskom meste). Nielenže sú schopné vyvinúť zaujímavo zložité správanie, ale vzájomne sa pozorujú a toto správanie sa replikuje z mozgu do mozgu. Majú teda vlastné mémosféry, len oveľa redšie ako máme my.
Replikácia mémbiontov formou pozorovania správania druhých
(extrémne dôležitá vec pre sociálne organizmy) je pomerne
neefektívny nástroj replikácie. Síce aj tu mémbiont-predok
vytvára zmeny v prostredí vo forme správania pozorovaného mozgom,
kam sa zreplikuje ako mémbiont-potomok, lenže takto vytvorené
zmeny v prostredí sú veľmi variabilné. Ak budeme tieto zmeny v
prostredí chápať ako formu génu, ktorý musí pozorujúci mozog
zreplikovať a zinterpretovať do vnútornej metabolickej štruktúry
mémbionta, tak tento gén veľmi variuje situáciu od situácie.
Napríklad tie vrany si nemôžu vždy zopakovať úplne presne rovnakú
situáciu na vozovke. Kdežto gén vo forme slova si veľmi ľahko pod
tlakom prírodného výberu ustáli svoju najpoužívanešiu formu.
Jeden z mojich známych má vnúčika, ktorý sa učí ešte hovoriť a nik nevie prečo si miesto slova traktor osvojil slovo kakoň. Lenže všetci okolo neho na ten istý objekt hovoria traktor, čo samozrejme v ňom vyvoláva spor. Nuž a postupne sa začalo objavovať aj uňho stále častejšie slovo tuaktou (stále ešte nevie vysloviť r). Prírodný výber negatívne potláča všetky iné varianty, ktoré sa vyskytnú, jednoducho preto, že slovo sa vždy a skoro každým dá vysloviť v jednej jedinej podobe, bez ohľadu na kontext, v ktorom sa vyslovuje. Nie je preto náhoda, že sa v nejakých mozgoch musel vyvinúť taký systém mémbiontov, ktoré mohli byť nezávislejšie od kontextu prostredia, ako mémbionty replikované pozorovaním správania iných hostiteľov. Nevieme akou zhodou okolností to nastalo u našich prapredkov, ale zjavne to nastalo, pretože jazykové štruktúry v našich mozgoch ich najviac odlišujú od iných primátov..
Akonáhle sa mohli mémbionty oslobodiť od svojich nositeľov do tej
miery, že mozgy sa museli stále viac a viac prispôsobovať
replikovaniu v nich obsiahnutých mémbiontov, stávali sa stále viac
závislé jeden od druhého a vytvárali vlastné ekostémy nad mozgami.
Z mozgov ako určujúceho prostredia pre ich život, sa stále viac
mozgy stávali pôdou pre replikáciu mémbiontov kultúry. Pre
kultúru, teda ekosystém mémbiontov, sú mozgy niečo ako pôda,
humus, v ktorom sa replikujú a v ktorom žijú.
Mimochodom, čo to vo svete mémbiontov značí, že žijú? Ako žijú?
Proteínové organizmy žijú tak, že zbierajú zdroje aby sa mohli
replikovať, prípadne potom investujú zdroje do potomkov, aby
vyrástli a mohli sa replikovať. Čo robia v mozgoch mémbionty?
Podľa mojej mienky sú pre mémbionty zdrojmi také metabolické
dráhy a cykly, ktoré udržujú ich intenciomnalitu a schopnosť sa
replikovať. Okrem vytvárania metabolických dráh a mémov samozrejme
v mozgu platí aj ich likvidovanie. Synapsie, ktoré nie sú aktívne
po čase neurón zlikviduje. Pre každého mémbionta je preto životne
dôležité udržiavať toky metabolických dráh, ktoré ho tvoria, v čo najčastejšom opakovaní. Lenže mozog je (ako som už viac krát spomenul)
reakčný systém. Metabolické dráhy sú spúšťané zmyslovými vnemami
alebo v cykloch defaultnej veľkoškálovej sieti (teda vtedy keď
akože rozmýšľame trebárs so zavretými očami). Z logiky veci je
zrejmé, že evolučnú výhodu majú mémbionty, ktoré dokážu do seba
zahrnúť pokiaľ možno čo najviac príležitostí na upevnenie svojich
synapsií. Zrejme to je možná podstata javu, ktorému hovoríme
vedomie. Vo vedomí neustále nejaký mémbiont uchvacuje metabolické
dráhy pozornosti (pozornosť je zložitý systém medzi hippocampom,
bazálnymi gangliami a thalamom). O tieto dráhy pravdepodobne
prebieha skutočný darwinistický boj, pretože vlastne pozornosť je
vždy iba jedna a tak o ňu treba kompetovať.
Pre pozornosť je dôležité vedieť zapájať metabolické dráhy významov pre organizmus, tvoriacich jeho intencionalitu. Teda napríklad sexuálne, boj o hierarchiu v tlupe, potravinové a podobne, dráhy, ktoré majú intencionálny význam pre hostiteľský organizmus. Také mémbionty, ktoré vedia vytvoriť metabolické dráhy cez vrodené motivačné metabolické dráhy majú väčšiu šancu uchvacovať pozornosť a majú kompetičnú výhodu. Výhodou nie je len väčšia šanca na prežiti v mozgu, ale aj väčšia šanca na replikciu mimo mozog, pretože k tomu aby mozog reagoval smerom von musí mať na to smerovanú pozornosť.
Mémbiont má tým väčšiu šancu prežiť v mozgu a mimo neho, čím viac
metabolických dráh tvorí jeho intencionalitu. Vzhľadom na to, že
v bohatom konektóme mozgu je pomerne zložité udržať svoju
intencionalitu (je ťažšie vytvárať klauzuru), zdá sa, že je lepšia
iná životná stratégia, a to vytvárať symbiózy s čo najviac inými
mémbiontami. Čím viac symbióz, tým viac príležitostí na usmernenie
metabolických dráh cez všetky zúčastnené symbionty, následne
bohatšie príležitosti k replikovaniu mimo mozog. Je to vidno na
príklade slov, ktoré sú súčasťou mnohých mémbiontov. Výsledkom je
veľmi zložitý egosystém vzájomne husto previazaných symbiotických
vzťahov. Napríklad ak si predstavíme jablko, tak nepochybne budú
súčasne aktivované obrazové mémbionty, chuťové, rad skúseností s
jablkom, trebárs aj mémbiont receptu na jablkový koláč, a
samozrejme písaný a hovorený mémbiont jablka. Celkove sa zdá, že
čím je mémbiont jednoduchší, tým ľahšie si svoju klauzuru udrží,
teda jeho stabilita je vysoká. Opôť dobrým príkladom je slovo.
Naopak, čím je mémbiont komplexnejší, tým je preň ťažšie udržať
vlastnú klauzuru, a teda častejšie variuje mozog od mozgu.
Mimochodom, v tom tkvie omyl memetikov, ktorí považujú mémy za
lamarkistické.
Taktiež sa dá zrejme povedať, že v mozgoch máme hierarchický egosystém, v ktorom schopnosť symbiotizácie určuje misto v hierarchii. Niečo podobné ako je v proteínovom živote potravinová hierarchia (všimnime si, že aj tu ide v podstate o hierarchiu metabolických tokov). Na samom spodku hierarchie sú jednoduché mémy, a z nich vytvárané (často iba dočasne) metabolické dráhy. Príkladom takýchto mémov môžu byť mémy na grupách neurónov v oblasti V1, ako som vyššie popísal. Vyššie v hierarchii môžu byť jednoduché mémbionty a metabolické cykly, ktoré sú schopné vlastnej intencionality. No a potom stále vyššie v hierarchii stále komplexnejšie mémbionty so schopnosťou replikácie mimo mozog a s vlastnou prepracovanou architektúrou symbiotických štruktúr, ako napríklad názory na niečo, ideológie, viery v niečo.
Čo ďalej?
Účelom doteraz uvedeného bolo predložiť argumenty na podporu
predstavy mémovej formy života. Ak ju akceptujeme, nevyhnutne
vyvstáva mohutný rad otázok. Aký ten život je konrétnejšie? Aké
pravidlá v ňom panujú? Je zrejmé, že vnútorný život v egosystémoch
je stále intenzívnejšie regulovaný a modifikovaný nadmozgovou
organizáciou, ktorú môžeme nazvať mémosystémom, alebo kultúrou.
Ako sa to deje, aké sú tu pravidlá a ako sa vyvinuli? Taktiež
treba podrobnejšie a hlbšie spoznať ako sa tvoria vnútorné
egosystémy pod tlakom mémosystému, a aké tu platia pravidlá.
Napoko stále je nejasná vnútorná štruktúra egosystémov a ako je
závislá od vrodených mémov.
Dnes existuje rad vied, ktoré sa z rôznych uhlov zaoberajú tým, čo sa deje v kultúre. Rôzne ekonomické, antropologické, politologické, lingvistické a čo ja viem aké ešte disciplíny. Všetky sa pokúšajú popisovať javy v kultúre akoby nezávisle od človeka, apriorne vychádzajúc pri tom z nejakej zamlčanej predstavy o človeku. Táto predstava je zásadne antropocentrická. Jej základom je koncept slobodnej vôle, hoci vedecky už bol dávno vyvrátený. Človek tu vystupuje ako slobodný jedinec, ktorý sa rozhoduje na základe svojich potrieb a názorov. Dôsledkom rozhodnutí všetkých týchto slobodných jedincov je z pohľadu týchto vied potom nejaká zákonitosť, ktorá spätne dopadá na týchto slobodných a autonómnych jedincov. Hoci tomu očividne takto nie je, viera v autonómnosť človeka nepustí.
Myslím, že je treba paradigmatická zmena. Rozhodovanie ľudí nie je autonómne, iba dáva každémnu odmenu v pocite autonómnosti. Rozhodovanie ľudí je vopred sformované budovaním egosystémov živých bytostí, ktoré nazývam mémbiontami. Ony v každom mozgu postupne od narodenia budujú egosystém, ktorý potom rozhoduje a tým rozhodovaním spoluvytvára mémosystém. Konštrukčným princípom je Darwinov prírodný výber. Naše rozhodovanie je dôsledkom nie našej intencionality, teda intencionality opice druhu Homo sapiens, ale intencionality mémbiontov. Javy, ktoré vyššie spomenuté vedy popisujú preto majú svoju detemináciu v zákonitostiach intencionality mémbiontov, tam ich treba hľadať. Nedáva celkom zmysel merať súčasný stav povedzme v ekonomike, bez skúmania toho ako sa v minulosti vyvinul, prečo sa tak vyvinul, ktoré organizmy sa na tom podieľali tak, že sformovali rzhodovanie súčasnej populácie jedincov. Mémbionty, ktoré dokážu efektívnejšie využívať vrodené hierarchizačné mémy majú vyššiu motivačnú silu ako nejaké veľmi komplexne teoreticky prepracované koncepcie. Samovražedný útočník je ekonomický nezmysel, ale mémologicky je jasne uchopiteľný a jednoznačne vysvetliteľný.
Je jasné, že keď ide o tak zásadný paradigmatický zvrat v
myslení, ako mémológia predstavuje, je vylúčené sa spoliehať na
to, že niekto bude sám schopný vyrobiť v jednom mozgu celý
ekosystém vied, ktoré by parciálne z nového pohľadu všetko
reinterpretovali. To je ako keby mal Newton vymyslieť kompletne
celú fyziku až po kvantovku. V prvom rade treba skutočne hľadať v
mozgu odpovede na to, či a do akej miery je možno v ňom všetko
interpretovať ako živý systém. Ja sa domnievam, že áno. Bude však
treba ešte veľa výskumu v oblasti neurovied a ideálne by bolo,
keby sa nejaká časť tohoto výskumu viedla z pohľadu novej
paradigmy. Teda v snahe hľadať metabolické systémy s vlastnou
intencionalitou, miesto toho, že sa hľadajú odpovede na otázky:
ktorá časť mozgu sa zapája do ktorej činnosti.
Všetko začína v mozgoch a v mozgoch to aj žije.
Identitárska plemenitba
Identitárska plemenitba
Pohlavia sú len dve.
Šľachtených sexuálnych
plemien môže byť trebárs aj 72
Tak ako v stredoveku bolo každé spoločenské hnutie podmienené nejakým výkladom Boha, teológiou, tak aj v novoveku sa všetko zdôvodňuje vedou. Miesto náboženských hnutí obsadili ideológie. Marxov vedecký komunizmus, Hitlerov (vlastne Himmlerov) vedecky zdôvodnený rasizmus a iné ľavicové ideológie prinútili obrovské masy ľudí k pohybom, radikálnym zmenám v ich životoch a veľké masové vymierania. Ideológie, ktoré sa opierajú o vedu majú za sebou najmasovejšie genocídy v evolučných dejinách hominidov.

|
Nezvyknem sa hodnotovo vyjadrovať, skôr mi je vlastné pokúšať sa javy vysvetľovať ako ich hodnotiť. Lenže o vedu sa opierajúce ideológie sú veľmi nebezpečný jav. Západná civilizácia má sklon upadať do vojen názorov a jedna takáto vojna sa rozhára aj v súčasnosti. Opäť vidíme nárast ideológií, ktoré svoje (z kontextu vytrhnuté) tvrdenia opierajú o vedecký výskum. Sú ideológie, kde sa veda spája s mocou. Snáď je ešte čas, kedy sa to spojenie podarí prerušiť.
Rozhodol som sa aktívne vystupovať proti ľavicovej identitárskej ideológii na poli, ktoré je mi najbližšie. Budem systematicky spochybňovať vedecký a filozofický pilier, na ktorom idenitárstvo stojí. Momentálne je jadrom identitárstva boj o sexuálnu identitu. Preto:
- Uvediem na pravú mieru hlavné interpretácie vedeckého výskumu, o ktorý sa v argumentovaní obhajcovia sexuálneho identitárstva opierajú.
- Budem dokazovať, že pre náš druh homosexualita mužov neprináša nijaký evolučný prospech.
- Prinesiem argumenty (opierajúce sa často o ten istý vedecký výskum) pre tvrdenie, že miera homosexuality je kultúrne, nie biologicky podmienená.
- Prinesiem argumenty pre tvrdenie, že homosexualita mužov je prírodným výberom v pôvodnom prostredí negatívne selektovaná a v kultúrnom prostredí môže byť pozitívne selektovaná.
- Budem obhajovať mémologickú hypotézu, že 72 pohlaví a iné formy zdanlivo absurdných identít sú prejavom tretej fázy domestikácie, teda plemenitby. Cielenej selekcie a úpravy mozgov kutúrou za účelom výroby nových plemien sapientov.
Kedže sa pravidelne stretávam s neschopnosťou najmä ľavičiarov čítať text s porozumením, tak explicitne hovorím, že toto moje úsilie nie je zamerané proti homosexuálom, ale proti identitárskej ideológii, ktorá v konečnom dôsledku škodí aj im.
Prečo je identitárska veda nepravdivá
Prečo je identitárska veda nepravdivá
Obráncovia LGBTI ideológie sa samozrejme v módnom trende opierajú o vedu ako o slovo Božie. Zdá sa mi, že ak cbce niekto dostať grant na vedecký výskum v psychológii, sociológii, či antropológii, tak má väčšiu šancu ak ho orientuje na témy, ktoré majú dokazovať prirodzenosť a normalitu sexuálnych deviácií, či problémov s vnímaním vlastnej identity.
Predtým, ako sa budem venovať ich argumentom, považujem za dôležité vysvetliť epistemologický rámec týchto vedeckých výskumov. Keď v 13. storočí františkánsky mních Roger Bacon formuloval presvedčenie, že Boha nemáme spoznávať v našich hlavách popletených predstavami, ale v jeho diele, v Prírode, vznikol jeden z pilierov modernej vedy — empirizmus. Nešpekulujeme prečo je niečo v prírode tak, či onak, ale dôkladne sformulujeme otázku (napríklad vo forme experimentu) a z odpovedí, ktoré nám príroda dá sa snažíme pochopiť podstatu javu, na ktorý sa pýtame.
Ako sa pýtaš, tak sa dozvieš
Je preto dôležité ako otázku položíme. Otázke je neoddeliteľnou súčasťou odpovede, ktorú nám príroda, či Božie dielo, dá. Do akej miery je otázka súčasťou odpovede veľmi závisí od zložitosti javu, ktorý skúmame a od toho, ako je oddelený ten čo sa pýta od javu, ktorému kladie otázku. Potom je niečo iné pýtať sa ako sa správa hmnotný bod v gravitačnom poli a niečo iné pýtať sa dotazníkom na sexuálne preferencie. Otázky na javy pôsobiace vo veľmi zložitých sieťových vzťahoch sú nevyhnutne redukcionistické, neschpopné zakomponovať celú nesmiernu zložitosť vzťahov, ktoré súčasne na jav pôsobia.
Navyše kladenie otázok v sociálnom prostredí súčasne to prostredie mení Podobne aj vo fyzike: čím viac sa experiment blíži ku kvantovým hodnotám, tým viac je otázka súčasťou odpovede. Predstava subjektu skúmajúceho objekty sveta mimo neho je principiálne nesprávna. Ako uvidíme v ďalších kapitolách, súčasný intenzívny záujem o sexuálne plemená [1] v Západnej civilizácii pôsobí cez aktivizmus rôznych skupín spätne na mentálny vývoj nových generácií a nevyhnutne mení výsledky javu, ktorý veda skúma. Otázka je tu výrazne formatívnou súčasťou odpovede.
Aby sme si rozumeli. Pýtať sa treba, veda je tu od toho. Treba si však vždy zachovať veľkú opatrnosť pri interpretovaní výsledkov výskumu ako definitívnej pravdy o svete, a to sa žiaľ v týchto otázkach veľmi často zanedbáva. Formulovanie otázky najmä v humanitných vedách je veľmi často zaťažené nejakým základným hodnotovým pohľadom vedca. Preto si pri čítaní rôznych článkov (aj odborných) na tému identity treba dať vždy tú prácu pokúsiť sa preformulovať otázku inak, z iného základného hodnotového postoja, a následne výsledky výskumu reinterpretovať. Ak to ide, tak máme dôvod k silnej nedôvere.
Veda neodhaľuje pravdu o svete
Súčasťou západného základného mentálneho nastavenia je predstava, že veda vie dokázať čo je pravda a čo nie. Vety ako „vedci dokázali…“ alebo „verím vede…“ sú úplne bežnou súčasťou argumentačného repertoáru v spoločenských diskusiách. Za tým je skrytý implicitný predpoklad, že ak niečo veda dokáže, tak je to hotová a večná pravda a každý názor, ktorý sa neopiera o vedu je vopred odsúdený k tomu byť omylom.
Český filozof Ivan Havel tento náboženský prístup k vede nazýva scientizmom (Vesmír, O scientizmu, Vesmír 76, 1997). Jeho základné axiómy odhalil v tejto štruktúre:
- veda odhaľuje pravdu,
- pravda je jediná,
- veda jediná odhaľuje pravdu,
- pravda je to, čo odhaľuje veda.
Ide samozrejme o sebapotvrdenie v dôkaze kruhom, niečo tak typické pre všetky náboženstvá. Aby sme vedeli povedať definitívne pravdy o svete, musel by byť svet súčasťou nás, nie my súčasťou sveta. Ak by sme ho celý obsiahli v sebe, tak by sme poznali všetky súvislosti, poznali by sme celú pravdu. S pravdou je to tak, buď je úplná, alebo nie je. Nie sme absolútne bytosti, máme preto iba možnosti, pravdepodobnosti, že svet je taký, či onaký.
Význam vedy je v jej metóde, ktorou dokáže kolektívnym úsilím vedcov eliminovať chyby vo vytváraní nášho obrazu sveta. Podstatou tejto metódy je principiálna nedôvera vo výsledky nášho poznania. Každé vedecké poznanie platí iba do tej chvíle, kým nie je vyvrátené. To značí, že mu treba nedôverovať, inak by platilo večne a malo by charakter náboženskej dogmy. Treba ho neustále spochybňovať, hľadať spôsobo akým ho vyvrátiť.
Interpretácia a jej kontext
Jedným z dôsledkov veľkej zložitosti živých systémov je, že právd o nich môže byť veľa, stačí zmeniť uhol pohľadu na ten istý vedecký výskum.
Existuje napríklad výskum, podľa ktorého ak majú matky viac synov, tak tí poslední majú vyššiu pravdepodobnosť, že budú homosexuálni. Nevieme celkom dobre prečo, ale najčastejšie sa spomína možnosť, že imunitný systém matky sa naučí útočiť po prvom synovi na vyvíjajúcisa plod s chromozómom Y. To hypoteticky posilňuje úlohu chromozómu X a chlapec sa vyvíja viac žensky. To slúži ako argument LGBTI+ aktivistom pre tvrdenie, že homosexualita mužov je prirodzená, normálna, veď inak by tomu prírodný výber zabránil. Formulujú hypotézy na čo je to nášmu druhu užitočné, ak časť mužskej populácie nemá potomkov ale zato môže prispievať k prežitiu potomkov súrodencov atď. Ako uvidíme v ďalšej časti tohoto cyklu. dá sa rovnako postaviť presne opačná hypotéza, a aj iné.
Ako je to možné? Ak môžeme vykladať tie isté poznatky vzájomne sa vylučujúcimi interpretáciami, je vôbec možné rozlíšiť, ktorá z nich je reálnejšia a ktorá menej? Pre formulovanie hypotéz či teórií sme si postupne osvojili niekoľko princípov, ktoré sa zdajú racionálne a sú všeobecne uznávané. Napríklad Occamova britva, ktorú možno parafrázovať do modernej podoby: z dvoch teórií je tá lepšia, ktorá vychádza z menej predpokladov. Alebo princíp falzifikácie, ktorý od tvorcu vedeckej teórie požaduje, aby ju formuloval tak, aby sa dala falzifikovať objavením nových dôkazov. Oba princípy sú často stúpencami identitárskych teórií ignorované.
Tu sa však zameriam na jeden podľa mojej mienky dôležitý princíp, a ten by sa dal formulovať: svoje tvrdenie, hypotézu, teóriu vždy udrž v celkovom kontexte dosiahnutého poznania. Máme tu napríklad vyššie spomínané tvrdenie o matkách, majúcich viac synov. Súčane však nemožno nevidieť, že v agránych spoločnostiach (kde sú viacerí synovia pravidlom a nie výnimkou) podiel homosexuálov nepresahuje mieru v iných kultúrach. Niečo tu potom nesedí, a ak niečo nesedí, treba byť veľmi opatrný s akoukoľvek interpretáciou spomínaného výskumu.
Obecne sa dá povedať, že čím širší je kontext poznania, o ktorý sa nejaký výskum opiera, tým sú jeho výsledky dôveryhodnejšie. Princíp falzifikácie nám naopak umožňuje menej dôverovať výskumom, ktoré sú v rozpore s inými výskumami. Obávam sa, že veľká časť identitárskych výskumov má len jeden kontext: dokázať fluidnosť našej prirodzenosti aby mohli platiť o predstavy o slobodnej voľbe toho kto som. Nech to stojí hoc aj celé dosiahnuté poznanie ľudstva.
- Pojmy sexuálne a identitárske plemená vysvetlím v samostatnej časti, kde sa opriem o teóriu domestikácie sapienta kultúrou.
Koľko je homosexuálov?
Koľko je homosexuálov?
Téma ochrany práv sexuálnych deviácií plní neustále verejný priestor. V pozadí verejnej diskusie prebieha diskusia odborná. Verejnej diskusii sa poddarilo vytvoriť dojem, že otázky okolo sexuálnej identity sa týkajú nejakej spoločensky významnej skupiny obyvateľstva, že ide o veľký spoločenský problém. Poďme sa teda najprv pozrieť aká veľká je tá skupina.

|
|
Percentuálne podiely sexuálnej orientácie podľa [1]
|
Údaje z rôznych zdrojov sa rôznia v rozsahu 1% gayovia a 1.5% lesby [2] po neuveriteľných 10% [3]. Na obrázku uvádzam graf z článku, ktorý sa primárne zaoberá nedostatočnosťou metodiky výskumu neheterosexuality. Ako z grafu vidno, umelo vytvorené kategóŕie ukazujú, že minimálne 94% opýtaných mužov a 86% žien sa jasne hlási k heterosexualite. Ktohovie ako by graf vyzeral, keby si mohli vybrať iba z krajných kategórií? Pretože ľudia (ako ešte bude reč inde) sa môžu pred dotazujúcim cítiť rôzne z rôznych dôvodov, a tie nemusia nutne súvisieť s pocitom sexuálnej identity.
Ako sa skoro vždy k týmto výsledkom výskumnici dopracovali? Správne tušíte — formou dotazníka. Dotazované osoby sa sami identifikujú kam patria. Sú aj iné, nepriame metódy, napríklad premietanie obrázkov a sledovanie kamerou očnú pupilu. Tieto sú však logisticky a finančne náročné a ako metóda sporné. Nikto vlastne nevie, či reálne niečo merajú.
Druhý problém je v skúmanej vzorke. Skoro výhradne ide o populácie, ktoré patria hlavne do civilizovaného prostredia Západu a ním kultúrne ovplyvneného Východu. Je ťažké si predstaviť, že bude výskumník pobehovať so svojím dotazníkom v Saudskej Arábii a očakávať, že nájde aspoň jedného homosexuála. Výskum v lovecko-zberačských spoločnostiach je prakticky nemožný a výskumníci narážajú hlavne na obrovské kultúrne rozdiely vo vnímaní sexuality aj v pojmoch a ich významoch.
Prečo zdôrazňujem kultúrne rozdiely? Výsledky totiž od nich závisia. Ak sa niekto cíti tak či onak, alebo to iba deklaruje, tak nijako nemožno vylúčiť, že homosexualita, či iná sexuálna deviácia je prinajmenšom jedným z dôsledkov kultúrneho selekčného tlaku na sapientov. Už vôbec nie je ako zistiť, či platí tvrdenie, že homosexualita je prirodzená. O prirodzenosti však bude reč podrobnejšie inokedy.
Tu si položíme otázku, ako sa prípadne vyvíjal počet sexuálnych deviantov počas histórie sapientov od paleolitu po dnes. To je dôležité vedieť, pretože ak to vieme nejako odhadnúť, tak vieme niečo podstatné o povahe tohoto javu. Po Darwinovi nemôžeme predpokladať, že máme nejakú vlastnosť, ktorá sa z ničoho nič objavila pred pár tisíc rokmi a dovtedy nebola. Preto musíme veriť, že neheterosexualita bola v našom druhu v nejakej podobe vždy. Lenže ako to zistiť? Z paleolitických vykopávok kostier to sotva zistíme. Aj keď získame zvyšky DNA, nevieme zistiť nič, pretože sa nepodarilo zatiaľ dokázať nejakú genetickú súvislosť medzi homosexualitou a konkrétnymi alelami génov.
Zostávajú iba nepriame indície. Napríklad pohľad na také skupiny sapientov, ktoré žijú snáď veľmi podobným spôsobom života ako naši prapredkovia, v podobných prostrediach, teda posledné zvyšky lovcov-zberačov. Pri prieskume k tejto téme som narazil na antropologickú štúdiu [4], ktorá opísala sexuálne zvyklosti lovcov-zberačov Aka a Nganda z centrálnej Afriky. Medzi inými stránkami sexuality sa výskumníci pýtali aj na homosexualitu. Príslušníci oboch kmeňov vôbec nechápali otázku. Nič také nepoznali. Načo by bol styk medzi ženami, či mužmi, však z toho by neboli deti?
Výsledky svojho výskumu porovnali s inými výskumami, hlavne s [5]. Táto štúdia porovnávala 20 parametrov sexuálneho správania zo Standard Cross Cultural Sample (SCCS), čo je najreprezentatívnejšia databáza rôznych parametrov správania zo 186 kultúr sveta. Podľa [5] 59% kultúr nepozná homosexualitu, tak ako Akovia a Nganovia. Značí to, že tam niet homosexuality?
Samozrejme, že sa s vysokou pravdepodobnosťou vyskytuje. Ibaže tlupa lovcov-zberačov má v priemere 80-120 členov. Koľko sa v nej vyskytne homosexuálov ak ich je v populácii povedzme 1%? Jeden? S kým bude mať vzťah? Podľa mňa je skôr pravdepodnejšie, že netuší čo s ním je a nejako neúspešne prežije svoj život. Pretože úspech sa meria deťmi. Je zrejmé, že sa preto u prírodných národov ťažko dozvieme aká je prirodzená miera homosexuality.
Môžeme však skúsiť urobiť odhad na základe nejakej civilizačnej postupnosti. Práve o to sa pokúsili autori štúdie [6], ktorá priniesla argument pre podporu hypotézy, že homosexualita mužov musí byť viac prevalentná v agrárnych spoločnostiach, pretože tam sa zväzky ženícha a nevesty dohadujú. Preto by tam mal byť pravdepodobne menší tlak negatívnej selekcie prírodného výberu. Dáta čerpsali z už spomínanej SCCS databázy.
Po štatistickom kúzlení im vyšlo, že oproti lovecko-zberačskej spoločnosti majú spoločnosti, ktoré sa živia chovom zvierat 2.7 krát, spoločnosti, ktoré sa živia zmiešaným poľnohospodárstvom 3.2 krát a lovom, zberačstvom spolu s chovom zvierat 1.6 krát vyššiu pravdepodobnosť, že bude v nich prítomná homosexualita. No a 6.4 krát vyššia pravdepodobnosť má byť v stratifikovaných agrárnych spoločnostiach. Ak by sme zobrali výsledky tohoto výskumu za platné, tak by bola prirodzená miera homosexuality hlboko pod 1%. Každé väčšie percento by potom bolo následkom pôsobenia kultúry, či priamo ako selekčný tlak, alebo nepriamo ako nedostatok selekčného tlaku prírodného výberu.
Záverom tejto kapitoly chcem znova zdôrazniť, že aj na tak jednoduchú otázku, akou je podiel sexuálnych deviácií na celkovej populácii, je neľahké zodpovedať. Živé systémy nie sú fyzikálne, neplatia v nich jednoduché kauzality. Nedá sa preto od výskumov očakávať, že prinesú jasné odpovede. O to horšie je, ak si v diskusiách vezme každá zúčastnená strana výsledky výskumu, ktorý jej vyhovujú a ignoruje tie výskumy, ktoré jej nevyhovejú. A to sa bežne v identitárskych sporoch deje.
-
Bailey, J. M., Vasey, P. L., Diamond, L. M., Breedlove, S. M., Vilain, E., & Epprecht, M. (2016). Sexual orientation, controversy, and science. Psychological Science in the Public Interest, 17(2), 45–101.
- Tento údaj mám z Is 10% of the population really gay?
Je to z britského ľavicového plátku Guardian. Problém tohoto zdroja je, že sa odvoláva na zdroje, ktoré sa mi nepodarilo dohľadať, pretože linky odkazovali niekam inam.
- Tento údaj pochádza z výskumov legendárneho sexuológa, ktorý vlastne sexuologický výskum založil, Alfreda Kinseya. Knihy: Sexual Behavior in the Human Male (1948) and Sexual Behavior in the Human Female (1953)
- Hewlett, B., & Hewlett, B. (2010). Sex and searching for children among Aka foragers and Ngandu farmers of Central Africa. African Study Monographs, 31.
- Broude, G. J., & Greene, S. J. (1976). Cross-Cultural Codes on Twenty Sexual Attitudes and Practices. Ethnology, 15(4), 409–429.
- Apostolou, M. (2017). Is Homosexuality more Prevalent in Agropastoral than in Hunting and Gathering Societies? Evidence from the Standard Cross-Cultural Sample. Adaptive Human Behavior and Physiology, 3.
Neverte na evolučný paradox
Neverte na evolučný paradox
V identitársky orientovanej vede sa často argumentuje s pojmom evolučný paradox [1]. Ten by sa dal formulovať asi takto: prírodný výber uprednostňuje také riešenia, ktoré vedú k čo najviac potomkom, homosexualita však pôsobí opačne, čo je vraj paradoxné. Najmä homosexualita mužov, pretože homosexuálne ženy predsa len môžu mať potomkov napriek svojej orientácii ľahšie.
Paradox však je iba v tom, že žiaden evolučný paradox nie je. Je to fixná idea, ktorú musia riešiť iba identitárski ideológovia po tom ako si ju stvorili.
Nepovažujú za evolučný paradox výskyt pedofilov, či zoofilov. Alebo schizofréniu, ktorá má podobnú prevalenciu v našej kultúre ako homosexualita a tiež znižuje zdatnosť (fitness) druhu. Identitárski ideológovia radi spomínajú, že evolučným paradoxom sa zaoberal už Darwin, len zabúdajú, že Darwin ešte vôbec nevedel ako sa dedia znaky, nepoznal genetické zákonitosti. Preň homosexualita naozaj mohla byť paradoxná.
Na to aby sme evolučný paradox akceptovali treba viac predpokladov ako ku tvrdeniu, že evolučne je všetko v poriadku. Treba uplatniť Occamovu britvu: hypotéza, ktorá vychádza z menej predpokladov je platnejšia ako taká, ktorá potrebuje predpokladov viac. Hypotéza, ktorá tvrdí, že aj v tomto prípade platí normálne prírodný výber vychádza z jedného predpokladu. Hypotéza, ktorá vychádza z toho, že platí prírodný výber plus homosexualita prináša evolučnú výhodu, potrebuje minimálne predpoklady dva. Obe hypotézy rovnako nevedia výskyt homosexuality kauzálne vysvetliť, o čom ešte bude reč.
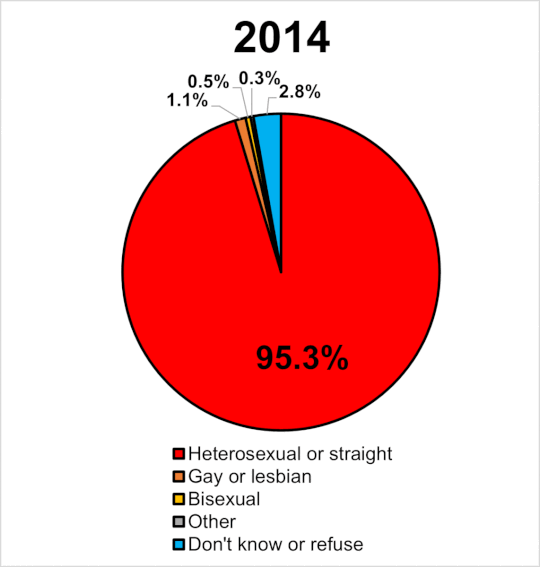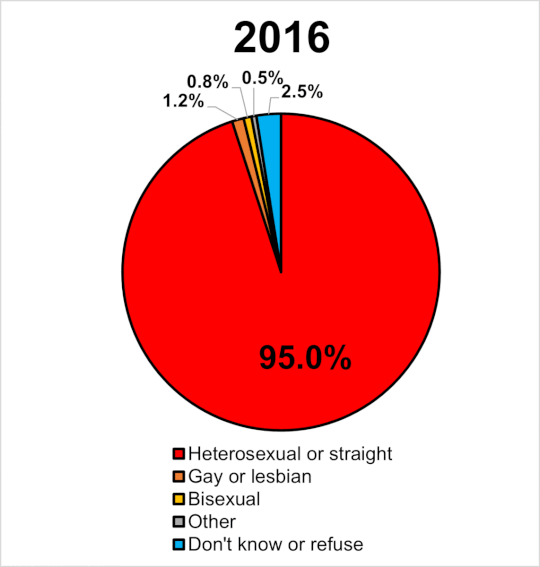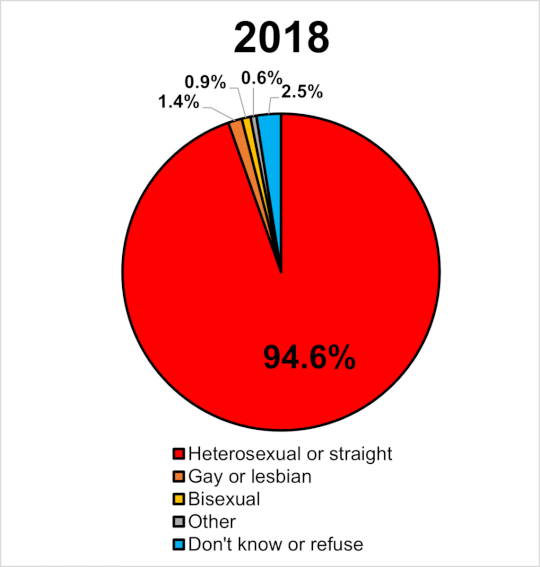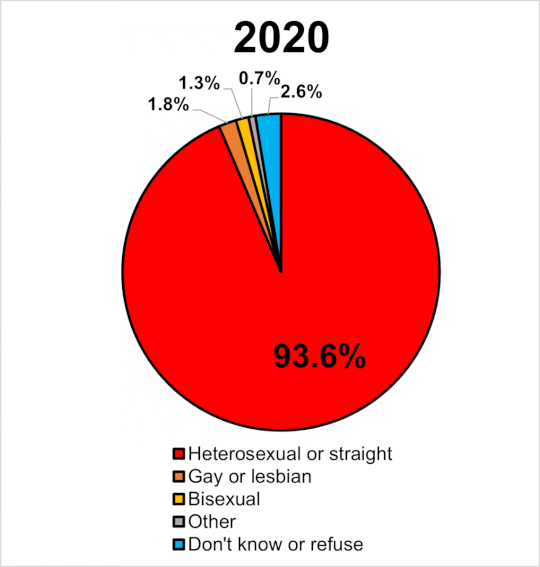
|
|
Miera sexuálnej orientácie v UK 2014-2020. Wikipedia
|
Podľa [2] (ktorý je citovaný v [1]) je homosexualita pomerne slušne dedivá, pohybuje sa dokoca v desiatkach percent príbuzných. Výskum je samozrejme robený v našej kultúre, ktorá je pomerne tolerantná k vybraným sexuálnym deviáciám. Napriek tomu (ako som uviedol v Koľko je homosexuálov? ) je rozumné predpokladať, že aj za týchto podmienok sa miera homosexuality pohybuje pod 1.5% u mužov a okolo 1% u žien. To je na dedivosť, uvedenú v [2], pomerne malé číslo.
Navyše, ak by bola tak vysoká dedivoať a nefungovala negatívna selekcia prírodného výberu, zrejme by časom narastal počet homosexuálov. Tí by tvorili významný podiel obyvateľstva a pozorovali by sme efekt evolučne stabilnej stratégie. Ak sa pozrieme na graf v pripojenom obrázku, tak v GB od roku 2014 do roku 2020 stúpol počet gayov a lesieb o 63%. Celkove sa ich podiel na populácii zvýšil z 1.1% na 1.8%. Len ak je naozaj pravda, že je homosexualita významne dedivá a súčasne ak platí, že nie je z populácie vyraďovaná negatívnou selekciou (ako tvrdia identitárski ideológovia) tak za tie tisícročia by sme to mali mať rtozhodne oveľa vúčší počet homosecuálov, ako pozorujeme.
Samotný výskum sa opiera o vágnu metodiku, čo konštatujú samotní autori. Napriek tomu prečo nie? Určite je nejaká dedivosť, ostatne ako u každého znaku. Ak sa naprieč mnohými kultúrami miera homosexuality udržuje v približne rovnakých číslach, tak to svedčí skôr o tom, že pôsobí negatívna selekcia prírodného výberu, nie pozitívna. Je jedno, že zatiaľ nepoznáme mechanizmus pôsobenia, očividne pôsobí.
My totiž vôbec nevieme čo sa na úrovni génov dedí, čo má na vývoj sexuality vplyv. Ide o multigénnyy faktor a je dobre možné, že ak sa výskyt nejakých alel génov zvýši nad určitú mieru, začne v populácii pôsobiť negatívna selekcia prostredníctvom iných génov, ktoré sa s tými „homosexuálnymi“ častejšie dostanú do interakcie a negatívne ich odselektujú z populácie.
Evolučný paradox nie je faktom, ktorý treba vysvetliť. Je to iba hypotéza, ktorá ešte len hľadá fakty, o ktoré by sa oprela. Ba čo viac, skôr ide o nepochopenie javov a ignorovanie iných faktov ako tých, ktoré vyhovujú predstave evolučného paradoxu. Evolúcia je neparadoxná.
- Barron, A. B., & Hare, B. (2020). Prosociality and a Sociosexual Hypothesis for the Evolution of Same-Sex Attraction in Humans. Frontiers in Psychology, 10, 484290.
- PILLARD, R. C., & BAILEY, J. M. (1998). Human Sexual Orientation Has a Heritable Component. Human Biology, 70(2), 347–365.
Bieda teórií pozitívnej selekcie homosexuality I. časť
Bieda teórií pozitívnej selekcie homosexuality I. časť
Aby som sa neposmieval slamenému strašiakovi, najprv definujem čo
ideológovia homosexuality (HS) predpokladajú a v čom sa hlboko mýlia. Budem
sa opierať o prehľadovú štúdiu [1], v ktorej autori používajú pojem same-sex sexual attraction (SSSA), hoci hlavne sa bude riešiť mužská HS. Preto aj ja budem rozlišovať v texte oba pojmy podľa toho, aký kontext zrovna autori využívajú, aj keď ho zamlčia. Ako uvidíme, súčasťou argumentácie autorov je pomerne svojské vykladanie citačných zdrojov tak, aby vyzneli „správne“ a vytvorili lekový kontext a atmosféru ich štúdie.
Ideológovia SSSA predpokladajú, že:
- HS nie je sexuálna deviácia, ale sa líši niečím podstatným od iných sexuálnych deviácií, napríklad pedofílie, nekrofílie, zoofílie atď. Tie podľa nich sú sexuálnymi chorobami, či deviáciami. Rozlišovacie kritériá sú podľa nich dve, z čoho to prvé je čisto etické. Homosexuáli svojou sexualitou nikomu neškodia na tozdiel od povedzme pedofilov. Etickými otázkami sa ja nebudem zaoberať, nie sú relevantné vo vecnej a racionálnj diskusii.
- Druhé rozlišovacie kritérium je údajne v tom, že HS je pre náš druh prospešná, užitočná, účelová, preto ju prírodný výber pozitívne selektuje, na rozdiel od iných deviácií.
Ako prvé treba pochopiteľne uviesť akej veľkej časti populácie sa SSSA a HS týka. V kapitole Neverte na evolučný paradox som doložil, že vôbec nie je jednoduché odpovedať na takúto otázku. Ani autori [1] na to priamo neodpovedajú, zato používajú pojmy ako „vo väčšine súdobých kultúr“ je HS „relatívne zriedkavá“, zato SSSA sa vyskytuje v pozoruhodnej „väčšej menšine populácie“. Dovolávajú sa pri tom dvoch prác. V prvej riešia otázku iba v USA populácii [2], čo akosi nepodporuje problematiku vo väčšine súdobých kultúr. Druhá práca [3] síce kladie podobnú otázku, ale jej hlavný cieľ je kritické posúdenie postupov štúdií, ktoré sa HS zaoberajú.
Autori [1] uvádzajú, že prinajmenšom jedna tretina sexuálnej orientácie je prisúditeľná aditívnym genetickým faktorom. Opierajú sa pri tom okrem iného aj o štúdiu [4]. Tam sa síce naozaj hovorí aj o tom, že sa podarilo identifikovať 5 lokusov na genofonde, ktoré súvisia s SSSA, lenže práve tento výsledok by mal autorov [1] znepokojovať, pretože prinajmenšom spochybňuje ich tézu, že ak by pôsobil prírodný výber proti HS či SSSA, tak by bol tento jav dávno odselektovaný a nepozorovali by sme ho. Problém však je, že u multigénnych faktorov sa nedajú z populácie vylúčiť alely génov, ak ich treba vylúčiť všetky. Prírodný výber tlačí na fenotyp, nie genotyp. Tým, že sa určitý fenotyp, napríklad gay, nerozmnoží nevzniká situácia, v ktorej zaniknú všetky gény, ktoré ho formovali. Tie sa roztrúsene môžu vyskytovať v jeho príbuzenstve, kde nespôsobia HS, pretože nepôsobia súčasne s inými pre HS potrebnými génmi. Multigénny faktor sexuálnej orientácie vysvetľuje nízky výskyt SSSA, či HS napriek negatívnej selekcii prírodného výberu spôsobenej nižším inclusive fitness homosexuálov. Autori [1] si vlastne citovaním štúdie strelili do nohy.
Poďme teraz k hlavnej téme o ktorú im ide, teda ako by mohol prírodný výber pozitívne selektovať HS.
Teória sexuálnej antagonistickej selekcie
Podľa tejto teórie by prírodný výber pozitívne selektoval mužskú HS ak by prinášala v celkovom súčte príbuzných viac potomkov. Teda, ak by gény, ktoré spôsobujú HS u mužov prinášli ženským príbuzným viac potomkov. Táto teŕia je podporená výskumami, podľa ktorých k tomu naozaj dochádza. V [1] uvádzajú viac teoretických modelov na podporu tejto teórie [5]. Ide však o matematické modely postavené na predpoklade, že ženy v príbuzenstve HS naozaj majú viac potomkov ako je mužská línia. Ako sami autori [1] uznávajú, pre toto tvrdenie je len slabá podpora vo vedeckej evidencii. Podľa [3] sú tieto výskumy založené na evidenciách západného civilizačného okruhu, a tam (ako vieme) majú ženy málo potomkov, natalita je pod 2. Treba na základe tak obmedzených súborov dát prejaviť veľkú štatistickú odvahu niečo dokazovať. Výskumy na lovecko-zberačských spoločnostich neexistujú a boli by určite zaujímavejšie, pretože tam je priemerná natalita žien 6. Za mňa je teória sexuálne antagonistickej selekcie mŕtvo narodené dieťa.
Teória príbuzenskej selekcie (štedrý strýko)
Teória príbuzenskej selekcie (TPS) je postavená na myšlienke výhodnosti HS pre prežitie viac potomkov v príbuzenstve. Aby to fungovalo musí prínos HS mužov pre inclusive fitness príbuzných prevyšovať stratu fitness vyplývajúcu z toho, že sa HS nerozmnoží.
V [1] citujú štúdiu [6], v ktorej sa zaoberajú možnými evolučnými adaptáciami vo forme HS v živočíšnom svete. Na konci štúdie je prehľadná tabuľka typov adaptácie, jej evolučných príčin a typického predstaviteľa. V prípade TPS sa ako typický predstaviteľ uvádza človek, hoci autori sami uznávajú, že je to podporené len veľmi slabou evidenciou. To ma zaujalo, pretože by bolo dobré vedieť, či vôbec nejaké zviera vykazuje správanie vyhovujúce TPS. Tak som začal hľadať nejaký príklad v zvieracej ríši. A skutočne, exemplárne sa uvádza Albatros laysanský. V štúdii [7] sa uvádza, že:
„Tridsaťjeden percent párov albatrosov laysanských na ostrove Oahu tvorili samice a celkový pomer pohlaví bol 59 % samíc v dôsledku prisťahovania samíc.
…
Páry samíc so samicami vyhniezdili menej mláďat ako páry samcov a samíc, ale bola to lepšia alternatíva ako nehniezdenie. Vo väčšine párov samica-samica, ktoré vychovali mláďa za viac ako 1 rok, bolo aspoň jedno mláďa geneticky príbuzné s každou samicou, čo svedčí o tom, že obe samice mali možnosť sa rozmnožovať. “
Na tomto príklade je vidno aké je ošidné oháňať sa argumentom adaptívnej prirodzenosti HS typu: však sú pre toto správanie príklady aj u iných zvierat ako sú ľudia.
Vtáky a opice sú si dosť taxonomicky a geneticky vzdialené. Je pomerne obtiažne (aj keď nie nemožné) predstaviť si taký genetický aparát, ktorý by rovnaké správanie udržal počas min. 70 miliónov rokov oddeleného vývoja. Skôr je pravdepodnejšie, že AK sa u oboch druhov takáto adaptácia objavila, tak ide o homopláziu, teda o rovnakú vlastnosť, ktorá bola nezávisle pozitívne selektovaná rovnakými podmienkami v prostredí. Známy príklad sú plutvy u rýb a vodných cicavcov (veľrýb, delfínov).
Práve o toto tu zrejme ide. HS správanie u ľudí a opíc je pozitívne selektované prostredím ak prináša rovnaký výsledok. Vidíme, že u albatrosa laysanského je nevyhnutná účasť dvoch dospelých jedincov na výchovu mláďat, takže ak z rôznych príčin je menej samcov, tak sa o výchovu starajú dve samice. To ovšem neznamená, že preferujú homosexuálne párenie. Hoci celkove vychovajú menej mláďat ako heterosexuálne páry, v konrečnom súčte to pre druh výjde lepšie. Tu naozaj teória príbuzenského výberu HS ako prirodzeného správania zrejme platí.
A čo človek? Výskyt jedincov, ktorí majú vyslovene homosexuálne preferencie je celkove veľmi malý, pod 2%. A to v najlepšie preskúmanej západnej kultúre. To je obrovský rozdiel oproti albatrosovi, kde také páry tvoria až 31%.
Keby išlo o homopláziu, tak by sme aj my mali mať túto vlastnosť v CELOM druhu, vo vštkých populáciách ľudí na svete, pretože by bola geneticky determinovaná. Problém však je, že na to nemáme vedeckú evidenciu. Dokonca niet evidencie, že v HS tolerantnejšej západnej kultúre by strýkovia a tety ochotnejšie podporovali svojich synovcov a netere, skôr naopak. Západná kultúra je silne individualistická a skôr potláča rodinné väzby ako by ich posilňovala.
Ako je to v iných kultúrnych prostrediach? U lovcov-zberačov nemáme reprezentatívnu evidenciu z dôvodov obtiažnosti takého výskumu. Podľa niektorých antropologických výskumov vo viacerých prírodných národoch HS ani nepoznajú.
Z agrárnych spoločností poznáme zvyky (kultúrny tlak), podľa ktorých sú príbuzní povinní postarať sa o synovcov a netere ak prídu o jedného alebo oboch rodičov. To však je iný prípad príbuzenskej selekcie ako cez HS. Je možné, že v niektorých kultúrach sa musia strýkovia a tety starať o príbuzných potomkov ak sú sami (potenciálne homosexuálni), ale štandard to rozhodne nie je.
V mnohých západných agrárnych spoločnostiach to v princípe tak bolo, len určite nebolo zvykom nemať vlastných potomkov. Stávalo sa, že najmä ženy ostávali „na ocot“, pretože sa ich nepodarilo rodičom vydať. Synovia spravidla tak nedopadli, pretože vždy so sebou prinášali nejakú pôdu alebo aspoň nejakú obživu, takže boli vyhľadávaní partneri. Ak boli homosexuáli, tak skôr boli bezdetní, ako by poskytovali výhody potomkom svojich súrodencov.
Predsa len mi však napadajú príklady aj v západnej spoločnosti, kedy kultúrne prostredie pozitívne selektovalo príbuzenský výber cez HS. Napríklad v anglickej viktoriánskej dobe sa stávalo, že veľa dievčat si nenašlo partnerov, pretože tí sa do ženby nehrnuli najmä z ekonomických dôvodov. Radšej viedli záhaľčivý život čo najdlhšie, neraz do konca života. Dokonca sa z toho medzi strednou vrstvou vytvorila dandyovská móda, ktorej reprezentantom je slávny spisovateľ Oscar Wilde, ktorý bol zrejme homosexuál, za čo bol aj odsúdený. Aby sa nejako morálne ospravedlnilo, že sa muž neožení, bolo veľmi rozšíreným pravidlom, že v živote podporoval deti svojich rôzne vzdialených príbuzných.
Podstatne rozšírenejšia forma TPS bola čisto kultúrne podmienená. Mám na mysli kresťanský zvyk krstných rodičov.
Z týchto príkladov možno vyvodiť jasný záver: ak sa u človeka vyskytuje príbuzenská selekcia cez HS, tak to NIE JE prirodzené správanie, ale správanie selektované na človeku kultúrou. Prírodný výber tu selektuje kultúrne normy, nie genotyp človeka. Možné to je iba preto, že fenotyp správania sa človeka je veľmi plastický vďaka vysokej komplexnosti mozgu.
(pokračovanie)
- Barron, A. B., & Hare, B. (2020). Prosociality and a Sociosexual Hypothesis for the Evolution of Same-Sex Attraction in Humans. Frontiers in Psychology, 10, 484290.
- Ward, B. W., Dahlhamer, J. M., Galinsky, A. M., and Joestl, S. S. (2014). Sexual orientation and health among US adults: national health interview survey, 2013. Natl. Health Stat. Rep. 15, 1–10
- Bailey, J. M., Vasey, P. L., Diamond, L. M., Breedlove, S. M., Vilain, E., and Epprecht, M. (2016). Sexual orientation, controversy, and science. Psychol. Sci. Public Interest 17, 45–101.
- Ganna, A., Verweij, K. J. H., Nivard, M. G., Maier, R., Wedow, R., Busch, A. S., Abdellaoui, A., Guo, S., Sathirapongsasuti, J. F., Team16, 23andMe Research, Lichtenstein, P., Lundström, S., Långström, N., Auton, A., Harris, K. M., Beecham, G. W., Martin, E. R., Sanders, A. R., Perry, J. R. B., … Zietsch, B. P. (2019). Large-scale GWAS reveals insights into the genetic architecture of same-sex sexual behavior. Science, 365(6456), eaat7693.
- Gavrilets, S., and Rice, W. R. (2006). Genetic models of homosexuality: generating testable predictions. Proc. Biol. Sci. 273, 3031–3038.
Camperio-Ciani, A., Cermelli, P., and Zanzotto, G. (2008). Sexually antagonistic selection in human male homosexuality. PLoS One 3:e2282. - Bailey, N. W., & Zuk, M. (2009). Same-sex sexual behavior and evolution. Trends in Ecology and Evolution, 24(8), 439–446.
- Young, L. C., Zaun, B. J., & VanderWerf, E. A. (2008). Successful same-sex pairing in Laysan albatross. Biology Letters, 4(4), 323.
Bieda teórií pozitívnej selekcie homosexuality II. časť
Bieda teórií pozitívnej selekcie homosexuality II. časť
Hlavným zámerom práce [1] je podľa autorov:
Navrhujeme sociosexuálnu hypotézu evolúcie SSSA, ktorá skúma možné adaptívne sociálne funkcie príťažlivosti a správania zameraného na rovnaké pohlavie.
Aby sa nejaké znaky v organizme vyselektovali ako adaptívne, musí na ne byť pozitívny selekčný tlak, ktorý ich zvýhodňuje:
… spájame evolúciu ľudskej SSSA so súborom znakov, ktoré sa vyvinuli ako dôsledok selekcie na ľahkú sociálnu integráciu (prosociálne správanie), toleranciu v rámci skupiny a sociálnu príslušnosť. Tento proces bol opísaný ako evolučný proces ľudskej autodomestikácie.
Autori vidia podporu pre svoju hypotézu v tzv. teórii autodomestikácie. Kedže sa otázkou domestikácie človeka zaoberám viac rokov, zaujímalo ma, odkiaľ berú argumenty pre svoju hypotézu. Tvrdia, že:
Sociálne evolučné procesy boli hlavnou hnacou silou súčasného ľudského poznania a správania, najmä selekcia na zvýšenie tolerancie v rámci skupiny a zníženie agresivity v rámci skupiny.
Možno to tak naozaj bolo, ibaže by to chcelo doložiť nejakými štúdiami. Na podporu svojho tvrdenia uvádzajú dve štúdie. V prvej [2] sa vôbec nezaoberajú našou evolúciou, ale skúmajú dlhodobý individuálny vývoj sledovaných jedincov v oblasti prosociálneho vývoja. V druhej [3] sa zaoberajú hypotézou, ako vzájomne na seba pôsobil vývoj sociality a kognitivity vo vzájomnom posilňovaní. Ani v tejto štúdii nenájdeme také silné tvrdenie o hlavnej hnacej sile vývoja nášho správania a poznania, ale o jednej z možných vývojových síl. Autori [1] často vynikajú až poetickými interpretáciami zdrojov a celkove mám dojem, že si z nich neprečítali poriadne ani zhrnutie (abstract).
Vráťme sa k hypotéze autodomestikácie, o ktorú sa autori [1] opierajú vo svojej argumentácii. Podľa [4] človek vykazuje tzv domestikačný syndróm, teda rovnaké znaky ako majú ním domestikované zvieratá. Domestikačný syndóm autori pripisujú selekčnému tlaku na prosocialitu, na také správanie, ktoré zvyšuje kooperatívnosť medzi členmi tlupy. To autori potrebujú k tomu, aby aj SSSA mohli zaradiť medzi prosociálne aktivity a tým odôvodnili pozitívny selekčný tlak aj na takéto správanie.
Problém však je, že je ťažké si predstaviť selekčný tlak na toľko znakov, ktoré zahŕňame pod jeden spoločný pojem prosociality. Oveľa pravdepodobnejšie vyzerá možnosť, že negatívnou selekciou na nejaký znak sa súčasne zmení správanie organizmu vo viacerých znakoch súčasne. No a to sa zrejme deje, ako preukázal svojimi experimentami s domestikáciou polárnej líšky D. K. Belyaev [5]. Pri jeho pokusoch sa selektovali z vrhov jedinci s najnižšou mierou agresie, ktorí boli pripustení ďalej do párenia. Po niekoľkých generáciách začali byť nielen menej agresívni (prosociálni), ale vykazovali celú škálu znakov domestikačného syndrómu, na ktoré experimentátori selekčný tlak nevykonávali.
Tu vidím slabé miesta argumentácie v prospech pozitívnej selekcie SSSA a v vlastne celej práce [1]. Selekčný tlak na pre druh neprirodzene nízku mieru agresivity zákonite „rozhasí“ normalitu jedincov a posunie ju do krajných polôh gaussovkej krivky. Napríklad jedinec má typický znak domestikačného syndrómu — juvenilný vzhľad a správanie — aj v dospelosti. V prostredí domestikácie sa stáva závislý od domestikátora, ktorý sa oň musí starať ako o mláďa. To by sa z pohľadu druhu mohlo označiť ako nevýhodný znak. Rovnako napríklad juvenilné hry, ktoré majú charakter SSSA, pretrvajú do dospelosti a to zideologizovaní autori označia ako pozitívny znak, ktorý je prosociálny, pretože im to vyhovuje.
Ďalej je samotný koncept autodomestikácie nanajvýš sporný. Nesporná a pozorovaná je domestikácia, kde domestikátor vytvára selekčný tlak na domestikanta na znaky, ktoré sú výhodné pre domestikátora. Domestikátor sa stáva prostredím domestikanta, pričom ho výrazne zbavil selekčného tlaku pôvodného prírodného prostredia. Autodomestikácia predpokladá, že tento proces si spraví druh sám sebe. Z nejakých hmlistých príčin začnú jedinci vzájomne potláčať agresivitu, následne sa vytvoria znaky domestikačného syndrómu, za čím nasleduje pozitívny selekčný tlak na znaky prosociality, napríklad aj na SSSA.
Otázka selekčného tlaku na potlačenie agresivity, ktorý by uprednostnil jedinca tak aby mal viac potomkov ako jeho agresívnejší sused ostáva nevyjasnená. Iba tak by sme totiž mohli hovoriť o seba-domestikácii druhu. V pohlavnom výbere uspievajú buď agresívnejší samci, či samice, alebo (ako u šimpanzov) samci, ktorí dokážu organizovať agresívne koalície voči iným samcom.
Máme vôbec nejakú evidenciu selekčného tlaku na potlačenie agresie v prírode? Vlajkovou loďou zastáncov autodomestikácie sú bonobovia, spolu so šimpanzmi nám najbližší opičí príbuzní. Skutočne, bonobovia vykazujú veľmi zvláštne prejavy správania, vrátane sexuálneho. Napríklad riešia konflikty vzájomnou masturbáciou, čím sa upokoja (toto majú priaznivci SSSA obzvlášť radi). Podľa pozorovaní je medzi samcami menej násilia a neboli pozorované prípady zabitia a násilia k samiciam [4]. Bonobovia navyše vykazujú viacero znakov domestikačného syndrómu, napríklad juvenilnejší vzhľad ako majú šimpanzy.
Problém s bonobami však je, že aj keď vidíme zvláštnu podobu ich správania, v skutočnosti púodľa viacerých pozorovaní sú dokonca v niektorých aspektoch správania agresívnejší ako šimpanzy. Citujem z abstraktu práce [6]:
Ako sa očakávalo, vzhľadom na to, že samice bežne prevyšujú samcov, zistili sme, že u bonobov je miera agresie samci-samice nižšia a miera agresie medzi samice-samce vyššia ako u šimpanzov. Prekvapivo sme zistili vyššiu mieru agresie samci-samci u bonobov ako u šimpanzov, aj keď sme analýzy obmedzili na kontaktnú agresiu. U oboch druhov dosiahli agresívnejšie samce vyšší úspech pri párení. Hoci naše zistenia naznačujú, že frekvencia agresie medzi samcami nie je paralelná s druhovými rozdielmi v jej intenzite, podporujú názor, že na rozdiel od šimpanzích samcov, ktorých reprodukčný úspech závisí od silných koalícií, samci bonobov majú viac individualistické reprodukčné stratégie
Takéto zistenie by som ja ako argument pre pozitívnu selekciu prosociality ako ju chápu v prácach [1, 4] nevidel. Skôr naopak. Ak kvôli vysokej samčej agresivite u šimpanzov sú šimpanzí samci schopní uzatvárať koalície, tak práve vysoká miera agresivity vyvoláva selekčný tlak na samčiu vyššiu schopnosť sociality. Bonobovia ostávajú evolučne pozadu v socialite. V štúdii [7] o tom aké je to prinajmenšom sporné hľadať vlastnosti človeka v našich opičích príbuzných sa tiež hovorí o agresivite bonobov:
Keď sa stretnú dve skupiny bonobov, často dochádza k agresii, pričom samce zvyšujú mieru agresie (Cheng et al. 2021). Na lokalite bonobov v Lomaku 35 % medziskupinových interakcií zahŕňalo fyzickú agresiu (Hohmann a Fruth 2002). Hoci v rezervácii Kokolopori Bonobo boli viackrát pozorované tolerantné medziskupinové interakcie, jedinci sa väčšinou socializovali s členmi vlastnej skupiny, nie s členmi cudzej skupiny, a 15 % stretnutí vyústilo do fyzických zranení (Cheng et al. 2022). V terénnej lokalite LuiKotale je tento model podobný: Počas trojmesačného obdobia intenzívneho výskumu došlo k 19 medziskupinovým stretnutiam medzi rôznymi skupinami bonobov, ale autori poznamenávajú, že medziskupinové stretnutia „boli viac agresívne ako tolerantné“ (Moscovice et al. 2022). Pri 47 % medziskupinových stretnutí došlo k „veľkoškálovým koaličným agresívnym udalostiam“, ktoré často viedli k zraneniam. „Počas jedného incidentu sa niekoľko samíc WBp [názov komunity] zameralo na samicu inej komunity May, a opakovane ju udreli a pohrýzli, keď sa snažila chrániť svoje malé dieťa“ (Moscovice et al. 2022).
Vysvetlenie odlišného správania bonobov a šimpanzov podľa tejto štúdie ostáva nejasné. Rozhodne však by som si ho netrúfal pripísať pozitívnemu selekčnému tlaku na „prosocialitu“, nech už to značí čokoľvek. Už vôbec by som na bonoboch nedemonštroval príklad autodomestikácie.
Vráťme sa k samotnému konceptu autodomestikácie. Vznikol už v 18. st. kedy si veľký prírodovedec Georges-Louis de Buffon všimol spoločných znakov domestikovaných zvierat a človeka a vyslovil názor, že aj človek je domestikovaný. Na rozdiel od iných domestikovaných zvierat, človek nemá nejakú divokú formu. Napríklad pes má vlka, koza divú kozu, atď. Vysvetlil si to tak, že niet človeka, ktorý by nežil v prostredí kultúry. Bol jeden z prvých, ktorí mal evolučný pohľad na živú prírodu a chápal, že znaky orgnizmov sú výsledkom pôsobenia prostredia. Ak človek pôsobí na zvieratá tak, že formuje ich znaky, na človeka takto pôsobí prostredie kultúry. Samozrejme antropocentrický názor človeka na seba ako na niečo výnimočné dáva logický záver. Kultúra je produktom človeka, a teda človek je jediné zviera, ktoré zdomestikovalo samé seba. Myšlienku autodomestikácie zopakoval o sto rokov aj Darwin a ujala sa najmä v nemeckej antropológii 30. rokov minulého storočia [8]. Znovu ožila v antropológii na konci tisícročia s obnoveným záujmom o náš domestikačný syndróm [9].
Chyba krásy však je v tom, že pojem domestikácie sa rozvíjal v prostredí antropológie. Ak sa na to isté pozrieme z pohľadu biológie, tak je zrejmé, že proces domestikácie je symbiotický proces, kde obaja symbionti vzájomne na seba pôsobia selekčným tlakom. Aspoň tak to zrejme bolo pred tisícročiami so psom a sapientom [10]. Jedinci domestikovaného druhu, ktorí boli menej agresívni, sa motali okolo ľuských sídiel a tí si možno spočiatku brali ich mláďatá na zábavu, neskôr na nejaký účel. Krížili ich medzi sebou a posilňovali takto domestikačný syndróm. Je očividné že selekcia bola na deviantných jedincov, pretože tí zrejme mali vo svojom prirodzenom prostredí kvôli nižšej schopnosti agresie selekčnú nevýhodu. V domestikácii preto nemôžeme hovoriť o selekčnom tlaku na prosocialitu, ale na nižšiu mieru agresie. To, čo autori [1] chápu ako prosocialita, je len kolaterálny efekt deviantnosti domestikovaných jedincov, ktorí zrazu môžu napriek svojej deviantnosti prežiť v prostredí, ktoré pre nich vytvorí domestikátor.
Jadrom symbiotického procesu domestikácie je intencionalita zúčastnených. Intencionálny záujem domestikátora o nejakú vlastnosť domestikanta je selektorom prírodného výberu. Domestikácia je v tomto špecifická forma symbiózy, pretože je v selekčnom tlaku asymetrická. Človek dokázal vyvíjať selekčný tlak na vlka, naopak to nešlo, aj keď nesporne nejaký selekčný tlak pôsobil. Vlk má zrejme takú genotypovú a fenotypovú plasticitu, ktorá v kontakte s človekom umožnila jeho domestikáciu.
Pre nás však je dôležité si uvedomiť, že aby tento symbiotický proces prebehol, tak je závislý od intencionálneho záujmu jedného zo zúčastnených. Preto je koncepcia autodomestikácie v jadre prinajmenšom sporná a podľa mňa vylúčiteľná. Je totiž ťažké si predstaviť aký intencionálny záujem by mali jedinci jedného druho voči sebe na znížení vzájomnej agresivity. Navyše keď vnútrodruhová agresia je výsledkom pochopiteľných evolučných tlakov najmä v pohlavnom výbere. Prečo by sa zrazu tento tlak mal stratiť? Domestikácia môže prebehnúť len medzidruhovo, nie v rámci druhu, pretože symbióza je možná iba medzi druhmi.
Autori [1] si zjavne neuvedomujú biologickú logiku domestikácie. Len im vyhovuje k tomu, aby zdôvodnili selekčný tlak na prosocialitu SSSA a v rámci toho aj HS. Dokonca jeden z autorov, Hare, v [4] pokojne zamení pojem domestikácie za autodomestikáciu aj v prípade domestikácie vlka. Podľa neho sa vlk sám domestikoval tak, že menej agresívni jedinci sa priblížili k ľuďom a tí ich adoptovali. Sami na seba vyvinuli selekčný tlak na prosocialitu a človek je v tom úplne pasívny. Homosexualita je jednoducho tým, za čo bola vždy považovaná: za sexuálnu deviácia, za patologické správanie, ktoré môže byť vedľajším dôsledkom domestikácie nášho druhu.
Ako ďalej uvidíme, na takto mylne chápanej domestikácii by malo byť založené evolučné zdôvodnenie homosexuality a homosexuálneho správania. Pri tom by sa aj SSSA aj HS dali vysvetliť ako dôsledok straty prirodzeného prostredia, nahradeného prostredím domestikátora. Domestikanti vykazujú rad patologických foriem správania (vrátane sexuálneho), ako svojimi pozorovaniami laboratórnych myší a potkanov popísal E. O. Price [11]. To dáva zmysel, na rozdiel od urputnej snahy hľadať nejaký komplikovaný evolučný mechanizmus pozitívnej selekcie, len aby sa dokaázala prirodzenosť neprirodzeného.
Otvorená ostáva otázka, že ak sa človek nedomestikoval sám, ako to, že javí znaky domestikačného syndrómu? Odpoveď na ňu však presahuje zámer tejto práce. Podrobne sa jej venujem na tomto webe v knihe
Principia memologiae.
- Barron, A. B., & Hare, B. (2020). Prosociality and a Sociosexual Hypothesis for the Evolution of Same-Sex Attraction in Humans. Frontiers in Psychology, 10, 484290.
- Eisenberg, N., Lennon, R., & Roth, K. (1983). Prosocial development: A longitudinal study. Developmental Psychology, 19(6), 846–855.
- dos Santos, M., & West, S. A. (2018). The coevolution of cooperation and cognition in humans. Proceedings of the Royal Society B: Biological Sciences, 285(1879), 20180723.
- Hare, B. (2017). Survival of the Friendliest: Homo sapiens Evolved via Selection for Prosociality. Annual Review of Psychology, 68(Volume 68, 2017), 155–186.
- Belyaev, D. K., Plyusnina, I. Z., & Trut, L. N. (1985). Domestication in the silver fox (Vulpes fulvus Desm): Changes in physiological boundaries of the sensitive period of primary socialization. Applied Animal Behaviour Science, 13(4), 359–370.
- Mouginot, M., Wilson, M. L., Desai, N., & Surbeck, M. (2024). Differences in expression of male aggression between wild bonobos and chimpanzees. Current Biology, 34(8), 1780-1785.e4.
- Glowacki, L. (2024). The controversial origins of war and peace: apes, foragers, and human evolution. Evolution and Human Behavior, 45(6), 106618.
- Stella, M. (2006). Teorie autodomestikace v německé antropologii a biologii do roku 1945. Kontext, důsledky, zastánci.
- Leach, H. M. (2003). Human Domestication Reconsidered. Current Anthropology, 44(3), 349–368.
- Komárek, S. (2012). Ochlupení bližní: Zvířata v kulturních kontextech. Academia.
- Price, E. O. (2003). Animal domestication and behaviour. CABI Publishing.
Mémologické zákonitosti konektómu mozgu
Mémologické zákonitosti konektómu mozgu
|
| Konektóm ľudského mozgu[3] |
Pre lepšie pochopenie významu mozgového konektómu treba priblížiť aká je architektúra mozgu. Jej základom sú populácie neurónov, ktoré sú vzájomne náhodne prepojené počas ontogenézy mozgu v maternici[1], [2] . Tieto populácie majú (podľa toho v ktorej časti mozgu sú) rôznu štruktúru, ale podstatné je vedieť, že vzájomne si v konkrétnej oblasti mozgu konkurujú v spracovaní rovnakého vstupného podnetu (napríklad v niektorom jadierku Thalamu, alebo v tzv. bunke primárneho vizuálneho kortexu). Podľa toho ako je populácia vnútri seba prepojená dokáže na tie isté podnety reagovať silnejšie alebo slabšie a túto reakciu odovzdajú ďalej do ostatných častí mozgu.
Mapu prepojení mozgu nazývame konektóm. Ide o akúsi schému prepojenia častí mozgu. Konektóm sa vytvára počas jeho ontogenézy v maternici. Jeho štruktúra je do značnej miery určená geneticky a epigeneticky. Štruktúra konektómu sa počas života vo veľkých škálach prakticky nemení, v stredných škálach viac a v mikroškálach najviac. Z toho pre mémbionty vyplýva zásadné obmedzenie vytvárania vlastných metabolických štruktúr, ale aj ekologických vzťahov.
Len pre doplnenie kontextu uvádzam, že prepojenia sú buď excitačné (posiňujú v cielených neurónoch reakciu), alebo inhibičné (reakciu tlmia):.
Z pohľadu mémológie je konektóm vrodenou metabolickou sieťou mémov. Z toho vyplývajú pre svet mémového života niektoré zákonitosti:
- Zákonitosť pozitívnej selekcie rastu komplexnosti konektómu
- Čím je konektóm komplexnejší, tým viac funkcií je mozog schopný vykonávať.
Konektóm mozgov cicavcov má počas evolúcie tendenciu mohutnieť. Jedinec s bohatším konektómom dokáže riešiť zložitejšie životné situácie, čo mu poskytuje evolučnú výhodu. Až od určitej hustoty konéktómov sú mémnbionty schopné replikácie do iných hláv.
- Zákonitosť komplexnosti metabolickej siete mémbiontov
- Čím je bohatší konektóm, tým môže byť metabolická sieť mémbiontov mohutnejšia a komplexnejšia
Čím je viac prepojení medzi časťami mozgu, tým je možné zapájať viac metabolických reťazcov do metabolickej siete mémbionta. Napríklad mémbiont slova láska, lokalizovaný prevažne v kortikálnej Brockovej oblasti, môže byť vnútorne regulovaný prepojením s podkôrovou amygdalou, ktorá vytvára pre sapienta emočný význam a pre mémbionta tohoto slova väčšiu šancu na replikáciu, pretože emočné slová sú častejšie replikované najmä ženskými mozgami.
- Zákonitosť bohatosti intencionality mémbiontov
- Čím je bohatší konektóm, tým môže byť bohatšia intencionalita mémbionta
S nárastom možností zapájania funkčných reakčných sietí do vlastnej metabolickej siete získava mémbiont viac možností zabezpečiť svoje životné záujmy, svoju intencionalitu.
- Zákonitosť komplexnosti ekosystému mémbiontov v mozgu
- Čím je bohatší konektóm, tým je schopnosť tvorby ekosystémových vzťahov mémbiontov väčšia.
Čím je konektóm bohatší, tým viac symbiotických a kompetitívnych vzťahov môže mať mémbiont. Čím je týchto vzťahov viac, tým sa zvyšuje jeho šanca replikácie do inej hlavy. Prírodný výber uprednostní mémbiontov, ktorí dokážu lepšie využiť konektóm.
- Zákonitosť pozitívnej selekcie plenárnych mémbiontov
- Čím je bohatší konektóm, tým majú na svoju evolúciu väčšiu šancu plenárne mémbionty.
Plenárne mémbionty sú také, ktoré veľkú časť svojej funkcionality prenechávajú na symbiotické vzťahy s inými mémbiontami. Tým je síce ich funkcionalita a štruktúra fluidnejšia, zároveň však sa zvyšuje vďaka tomu šanca plenárneho mémbionta na rozmnoženie. Z logiky veci vyplýva, že čím je komplexnejší konektóm, tým je väčšia možnosť plenárneho mémbionta vytvárať funkčné symbiózy s inými mémbiontami.
Zdroje:
- Edelman, G. M., & Rogers D. Spotswood Collection. (1992). Bright air, brilliant fire : on the matter of the mind. BasicBooks.
- Kandel, E. R. (2013). Principles of neural science Fifth edition (5th, rev. ed.). McGraw Hill.
- Wikipedia, Connectome, https://en.wikipedia.org/wiki/Connectome
Tri plus jedna otázky
Tri plus jedna otázky

Po mnohých rokoch pokusu o sformovania univerzálnej teórie mysle a kultúry sa postupne celá problematika sformovala do troch otázok. Tieto otázky vyvstali z nahromadenia obrovského množstva vedeckej evidencie za ostatných 400 rokov a všetky sa týkajú človeka, sú to teda antropologické otázky. Po celé obdobie vývoja našej kultúry tieto otázky neboli postavené z jednoduchého dôvodu, a to, že človek sám seba považuje prirodzene za centrum jeho diania. Lenže vedecký výskum nás postupne doviedol k inému obrazu, a toho dôsledkom je, že všetky humanitné, spoločenské, ekonomické, či antropologické vedy, ktoré sa pokúšajú vysvetliť človeka, ak nie sú schopné na tieto otázky dať konzistentnú odpoveď, tak určite nehovoria o človeku v súlade s dnešným stavom poznania špeciálnych vied. Určite sa preto často mýlia.
Tu sú:
1. Čo vyplýva z toho, že je človek domestikovaný organizmus?
Vedecká evidencia k tomuto je už dnes nespochybniteľná. Lenže kto človeka domestikuje? Biológovia hovoria, že domestikácia je symbiotický jav, čiže vyvstáva otázka, s kým žijeme v symbióze a prečo? Ako sa to stalo? Predstava tzv autodomestikácie je ľahko spochybniteľná a môžeme sa jej držať iba ak spochybníme podstatnú časť biológie.
2. Čo vyplýva z toho, že je myseľ deterministická?
Predstava slobodnej vôle bola odjakživa veľmi ťažko obhájiteľná. Darwinistický model evolúcie, dnes všeobecne prijímaný, túto možnosť priamio vylučuje. Vedecká evidencia v oblasti výskumu mozgu najmä od polovice 20. stor. postupne stále viac a viac odhaľuje síce veľmi ťažko popisovateľný, predsa však vo svojich princípoch deterministický proces, ktorý v našich mozgoch prebieha. Ktoré zákonitosti tam panujú? Ako sa vyvinuli a prečo?
3. Čo vyplýva z toho, že kultúra je evidentne darwinistický evolučný systém?
Dnes existuje bezpočet výskumov, ktoré kultúrne javy chápu ako evolvujúce. Všetky dnešné kultúrne elementy sa dajú zaraďovať do evolučných stromov. Kultúra ako celok je nevysvetliteľná bez toho, aby jej bola priznaná vlastná intencionalita, teda vlastnosť živého. Čo v skutočnosti teda v kultúre evolvuje a prečo? Odkiaľ sa to vzalo?
Všetky tri otázky majú zjednotiteľa v tom, že spolu súvisia v existencii druhu Homo sapiens, teda sa spájajú v človeku. Je tu teda logicky jedna zhrňujúca otázka:
4. Čo z toho všetkého vyplýva dokopy?
Teda inak povedané: potrebujeme nejaký nový koncept človeka, dôsledne neantropocentrický. Verím, že mémológia je schopná podať ucelený, vnútorne neprotirečivý, konzistentný model, ktorý odpovie na položené otázky jednotným spôsobom.
O živote v našich mozgoch
O živote v našich mozgoch
Tento článok vznikol ako ako odpoveď na úvahu chate na FB. Kedže sa rozrástla do nechatových rozmerov, tak som sa s FB priateľom dohodol, že ju uverejním ako článok.
Prosim ta, snazim sa predstavit ako vyzera prenos funkcie jednotlivca na skupinu. Napr. delba prace preniesla vyrobny proces niecoho z 1 cloveka na skupinu ludi, spolocnost.
Ak sa to ma preniest, tak musi existovat nejaky konsenzus medzi ucastnikmi tej skupiny, kto co bude robit, a musia vsetci suhlasit, a nejako sa koordinovat.
Tento konsenzus je nieco emergentne, nieco co tu je, ale predtym nebolo.
Bez konsenzu, to stale ostane v rukach jednotlivca.
Slovo konsenzus mi pride aj ako synonymum slova kultura.
No a takto sa v kulture nahromaduju tie vlastnosti jednotlivcov.
Mohol by som povedat ze vlastnost / funkcia jednotlivca sa rozdistribuuje na skupinu?
Cim je to vyhodne?
Tak jednotlivec sa zaspecializuje, urobi tu uzku cast lepsie, a vysledny produkt je kvalitnejsi, a pre vsetkych.
Ak sa to ma preniest, tak musi existovat nejaky konsenzus medzi ucastnikmi tej skupiny, kto co bude robit, a musia vsetci suhlasit, a nejako sa koordinovat.
Tento konsenzus je nieco emergentne, nieco co tu je, ale predtym nebolo.
Bez konsenzu, to stale ostane v rukach jednotlivca.
Slovo konsenzus mi pride aj ako synonymum slova kultura.
No a takto sa v kulture nahromaduju tie vlastnosti jednotlivcov.
Mohol by som povedat ze vlastnost / funkcia jednotlivca sa rozdistribuuje na skupinu?
Cim je to vyhodne?
Tak jednotlivec sa zaspecializuje, urobi tu uzku cast lepsie, a vysledny produkt je kvalitnejsi, a pre vsetkych.
Jadrom tohoto citátu v skutočnosti je otázka: odkiaľ a ako sa berú novinky? Odpoveď vôbec nie je jednoduchá, či jasná. Myslím, že v konečnom dôsledku je to jedna z najstarších a najnezodpovedanejších otázok ľudstva. Tvorí napríklad jadro vyše 2300 ročného sporu medzi Platónom a Parmenidom.
Ja môžem k nej povedať iba svoj názor, založený na mojich apriorných predpokladoch darwinistického konceptu sveta. Iba zopakujem, že nie som z tých, že by tento koncept považovali za pravdivý a za tú pravdu boli ochotní položiť život. Pravda je subjektívna kategória. Na druhej strane je tento koncept určite MOŽNÝ a ako taký je NEVYVRÁTITEĽNÝ. Téma je to obsiahla, preto som si na ňu musel nájsť čas. Takže k veci.
V darwinistickom svete sa objavujú novinky mutáciou predchádzajúcej generácie entít. Čiže jediným miestom v darwinovskom cykle, kde sa novinky môžu objaviť je moment replikácie. Následne sa tieto zmeny konfrontujú stávajúcim svetom, ktorý je výsledkom interakcií pred replikáciou týchto entít. Nastáva prírodný výber — ustanovenie novej ekosystémovej stability. Toto je pre túto tému kľúčové.
Kedže vlastne nepoznáme nijaký iný formujúci princíp, ako je ten darwinistický, odpovede musíme hľadať v ňom. A jednou z odpovedí je, že z darwinistického sveta nemôže vyrásť iný formujúci princíp ako opäť darwinistický. Prečo? Pretože v zásade máme tri možné formy mutácie:
- pozitívna (entita získava výhodu v prírodnom výbere),
- negatívna (entita je v prírodnom výbere znevýhodnená) a
- neutrálna (netreba vysvetliť).
Jedine o neutrálnej mutácii má zmysel uvažovať, ak hľadáme nejaký nedarwinistický formujúci princíp. Ako si to však v darwinistickom svete, ktorý stojí a padá na obmedzení zdrojov predstaviť? V ňom v skutočnosti nemôže byť neutrálna mutácia, pretože prinajmenšom zaťažuje entity energeticky replikovaním neutrálnych znakov, čím ich znevýhodňuje. Aby táto neutrálna mutácia uspela, musí nakoniec priniesť nejakú výhodu, ktorou tú energetickú záťaž vyvažuje. Ja skrátka na neutrálne mutácie neverím, aj keď existuje mnoho príkladov, ktoré sa na niečo také podobajú. Zabúda sa pri nich, že ich neutralita je len dočasná, že tzv. neutrálne znaky sú neutrálne iba v obmedzenom časovom rámci, že možno iba dočasne stratili svoj selekčný význam.
Pripomínam to preto, že práve v tej akože od prírody nezávislej zmene v mozgoch sa antropocentricky hľadá naša výhoda, ktorá nám udajne pomohla vymaniť sa z diktátu prírodného výberu. Nezmyselnosť tohoto konštruktu je obsiahnutý v jeho vnútornom paradoxe, pretože ako mohla nejaká neutrálna mutácia priniesť evolučnú výhodu vymanenia sa z diktátu prírodného výberu?
Nie, ak chceme pochopiť napríklad to, ako sa uskutoční „prenos funkcie jednotlivca na skupinu,“ tak musíme otázku formulovať darwinisticky: aká mutácia nejakej entity v darwinovskom svete umožnila preusporiadanie skupiny iných darwinistických entít (v tomto prípade ľudí)? A čo je vlastne tá entita, ktorá mutovala?
Ak chceme zodpovedať tieto otázky, tak si v prvom rade musíme uvedomiť, že skupina iba ťažko môže byť darwinistickou entitou jednak preto, že skupina sa nereplikuje, replikujú sa jedinci, nemôže preto mať svoju intencionalitu, nemá svoj metabolizmus a iba ťažko svoju klauzuru, teda štyri charakteristické črty živého. Nemožno preto preniesť z jedinca na skupinu nejakú životnú funkciu. Čo sa však dá je to, že skupinové správanie sa vyprodukuje ako VÝSLEDNICA všetkých individuálnych akcií živých entít, teda to, čo pomenúvame ako ekosystém.
Jedna z možností je, že sa napríklad mutáciou zmení vlastnosť jedincov druhu tak, že sa vytvorí určité kolektívne správanie. Takým príkladom je roj, či kŕdeľ. Roj sleďov ktorý sa pri ohrození predátorom zomkne a začne rotovať znemožňuje predátorovi zamerať sa na jedného sleďa. Také skupiny sleďov, kde sa miesto úteku mutovalo správanie na zomknutie mali väčšiu šancu na prežitie, ako ich predkovia, pretože umožnli tvorbu veľkých rojov, a teda väčšieho preživšieho množstva potomkov. Tu sa skupinová novinka objavila prostredníctvom mutácie jedincov toho istého druhu, čo vcelku dobre zodpovedá tradičnej darwinistickej predstave.
Horšie je to ovšem so symbiózami, kde tradičný darwinizmus veru nemá príliš plauzibilné vysvetlenie. Ako môže vzniknúť symbiotické spojenie medzi tak odlišnými druhmi, akým je trebárs riasa a huba v symbióze lišajníka? Potiaž je tototiž v tom, že prírodný výber sa opiera do synchrónnych mutácií v dvoch druhoch súčasne, čo je naozaj len veľmi ťažko predstaviteľné. Lenže tak tomu je a dokonca mnohí biológovia tvrdia, že symbiózy boli hlavnou hybnou silou evolúcie, pretože z pozície klasického darwinizmu sa iba veľmi obtiažne obhajuje nárast komplexity živých systémov vnútrodruhovými mutáciami.

Každý organizmus (teda tá darwinistická entita) nežije sama v sebe ale žije v prostredí, z ktorého je síce vyčlenená (to je tá Markošovská klauzura[1]), ale súčasne je doň včlenená, pretože potrebuje z prostredia zdroje, musí sa chrániť pred inými organizmami, atď. Pre svoj život, teda musí mať nejaké VEDENIE o prostredí. Kedže je vďaka klauzure, vyčleneniu, obmedzený v tomto vedení, tak sa jeho umwelt skladá z obmedzeného množstva ZNAKOV prostredia, ktoré sa v jeho innenwelte interpretujú v jeho intencionálnom VÝZNAME.
Ako príklad dobre poslúži dnes už notoricky uvádzaný Uexküllov kliešť. Kliešť má receptor na kyselinu maslovú, ktorá je súčasťou potu teplokrvných cicavcov (zdroj potravy) a tepelný receptor, ktorý mu umožňuje nájsť blizku žilku pod povrchom kože. Dva znaky — teplo a kyselina maslová — mu stačia pre to, čo potrebuje vedieť o svete, ak má jesť. Tieto znaky prostredia (umweltu) sa v jeho vnútri menia na jemu prospešný význam (innenwelt) fyziologických reakcií, metabolických kaskád, svalových sťahov, atď.
Spojenie s Darwinom je v tom, že ak rozdelíme znaky organizmu podľa von Uexkülla na znaky umweltu a znaky innenweltu, ktoré spolu súvisia, ale nutne nemusia jeden z druhého vychádzať, skôr sú väčšinou v nejakom flexibilnom vzťahu, tak vidíme pôsobenie selekčného tlaku v dvoch rôznych svetoch každej entity. Napríklad znak umweltu má určité flexibilné parametre (môže mať určitú tvarovú flexibilitu), pričom vyvolá rovnaké reakcie v innenwelte. Povedzme tvar postavy samice nášho druhu nemusí byť ideálny, ale stačí, ak sa okolo toho ideálu v určitom rozpätí pohybuje, aby jej šanca v pohlavnom výbere bola stále zachovaná, teda aby tento znak v innenwelte samca vyvolal rovnaký význam, teda vhodnosť k páreniu.
To, že prostredie pôsobí na entitu cez znaky jej umweltu je kľúčová úvaha tejto koncepcie. Prostredie je tvorené entitami, z ktorých každá rozoznáva ostatné entity pomocou znakov, ktoré má vo svojom umwelte. Komplexnejšie celky, také ako sú tie symbiotické, vzniknú potom z pohľadu prostredia tak, že nejaké pôvodne oddelené znaky symbiontov z pohľadu ostatných entít splývajú v jeden znak, voči ktorému majú entity prostredia už prirpravenú nejakú interpretáciu vo vlastnom innenwelte.
Napríklad ak krab pustovník naloží na seba sasanku, tak sa stáva pre okolité prostredie nepoživateľný, pretože sasanka jedovato pŕhli. Krab získal vlastnosť tým že spojil svoj umwelt s umweltom sasanky. Dôležité je si uvedomiť, že vznikol nový znak, znak symbiotického spolužitia, na ktorý pôsobí pozitívnou selekciou prírodný výber. Je to znak spojeného umweltu, nie znak umweltov, či innenweltov oboch účastníkov symbiózy. Až následne sa prírodný výber môže upriamiť na znaky innenweltov oboch účastníkov povedzme tak, že krab, ktorý sa mutáciami dopracuje k schopnosti rozoznať sasanky od iných objektov prostredia má väčšiu šancu na spoluvytvorenie symbiotického umweltu ako iní krabi a sasanka ktorá sa naučí rozoznať kraba a neútočí naň svojimi pŕhlivými zakončeniami má rovnako väčšiu šancu na prežitie, pretože vďaka pohybu kraba sa dostane k viac potrave.
Až následne môže nastať ŠPECIALIZÁCIA, teda prebudovanie innenweltov oboch symbiontov. Tak napríklad stačí črevnému parazitovi oveľa menej znakov jeho umweltu, aby prežil, čo následne vedie k jeho špecializácii na hostiteľa, a to následne vedie k radikálnemu zredukovaniu jeho innenweltu. Takýto stav redukovania innenweltu a umweltu môže viesť až k endosymbióze, kde povedzme u mitochondrií sa vlastne o nejakom umwelte ani nedá hovoriť.
Čo z toho vlastne vyplýva pre našu tému? Vznik tak komplexného kolektívneho správania, ako je to sapientie sa NEMOHLO vyvinúť mutáciou vnútri druhu, pretože takto sa môže vyvinúť nanajvýš správanie také, aké majú iné sociálne druhy, nejaký ten analogon správania kŕdľa sleďov. Naše mozgy sa od mozgov iných opíc líšia iba nepatrne, pričom s funkcionálneho pohľadu sú vlastne rovnaké. Komplexita teda určite nevznikla zvnútra nášho druhu. Takáto komplexita musí byť teda medzidruhová, musela sa vyvinúť selekčným tlakom na splynutie umweltov prinajmenšom dvoch druhov ŽIVÝCH entít.
Takže otázka na to, ktoré entity mutovali aby vytvorili napríklad spoločenskú deľbu práce nie je otázkou na mutáciu nášho druhu, ale nejakého iného. Tento druh nevzniká rozmnožovaním opice Homo sapiens, pretože táto sa rodí bez vlastností tej komplexity, ktorú nazývame kultúrou. Jediné vysvetlenie pre iný druh ako je ten náš, druh s ktorým žijeme v symbióze je, že tento druh nás osídľuje podobne, ako nás osídľujú rôzne druhy baktérií, s ktorými žijeme tak isto v symbiózach. Kým baktérie osídľujú naše črevá, dutiny, či povrch pokožky, tak ten druh, ktorý umožňuje vysokú komplexitu nášho správania osídľuje naše hlavy. Iná darwinistická možnosť vlastne ani neexistuje.
Dôležité je si uvedomiť, že tento druh nie je molekulárny druh, je to život podstane inej povahy, ale je to život, pretože musí mať vlastnosti živého — replikácia, intencionalita, klauzura a metabolizmus — aby mohol tvoriť symbiózy s naším druhom.
Naše mozgy sú osídlené mnohými živými bytosťami, a tie majú tak svoje innenwelty ako umwelty. A ich umwelty dokážu splývať a vytvárať symbiózy medzi sebou, čím vznikajú tie komplexity, ktoré by sme radi pripisovali vlastnostiam nášho druhu, hoci náš druh tieto schopnosti dokázateľne nemá.
Niet teda prenosu funkcie jednotlivca na skupinu, ale sú entity, ktoré vytvárajú také prostredie v našich mozgoch, ktoré umožňuje skupinové správanie. Ako? Tak ako každý iný formatívny akt v darwinovskom svete — prírodným výberom, tlakom na mutácie znakov živých entít v našich mozgoch. Deľba práce nevznikla, pretože to bolo výhodné pre každú opicu, čo sa na nej podieľala. Nič také pre opice nie je výhodné. Predsa je dokázané, že niet dobrovoľnej spolupráce v darwinovskom svete, a tie formy spolupráce, ktoré sa nám takými javia, sú také iba zdanlivo, viď napríklad recipročný altruizmus. V darwinovskom svete, kde každý jedinec je jedinečná skladba znakov a bojuje vlastne za zachovanie tejto skladby najviac ako sa len dá, tak deľba práce je príležitosťou pre parazitické správanie, presadenie vlastného genofondu na úkor druhého. Zrejme preto sa to nikde v prírode nevyvinulo, iba u nás a u sociálneho hmyzu, kde to však má úplne iný dôvod, kedže všetci členovia spoločenstva majú rovnakú DNA, sú to sestry, ktoré nemajú dôvod vzájomne sa podvádzať. Mozgy opíc musia byť k spolupráci DONÚTENÉ a prípadná výhodnosť takéhoto donútenia aj pre ich fitness je až sekundárna vlastnosť tohoto evolučného procesu, nie primárna.
Nikto si nesadol a nepovedal: pozrite, Fero robí lepšie šípy, Jano luky, Ďuro tetivy, tak čo keby sme si to rozdelili a všetci budeme mať najlepšiu výzbroj, no nie je to múdry nápad? Ak sa pozrieme na zozbierané antropologické štúdie dosiaľ fungujúcich lovecko-zberačských populácií, tak žiadna nevykazuje deľbu práce[2]. Každý robí všetko a súperí z druhými o svoj reprodukčný úspech.
Je teda zrejmé, že k špecializácii muselo dôjsť až po usadení sa na pôde po dlhom období samostatnej evolúcie živých bytostí v sapientích mozgoch. Tá deľba práce je ich vynález, je to spôsob ich, nie našich, symbiotických vzťahov.
Možno dokonca bol spočiatku veľmi nevýhodný pre sapientov podobne ako bol pre sapientov nevýhodný poľnohospodársky usadlý spôsob života. Je dokázané, že populácie, ktoré sa usadili na pôde trpeli mnohými chorobami, mali nižší vzrast a horší chrup. Kosili ich epidémie, ktoré v oddelených lovecko-zberačských tlupách nemajú až takú šancu. Väčšinou hladovali a umierali (na rozdiel od lovcov-zberačov) na vlny hladomorov z neúrod. Usadlý spôsob života sa napriek tomu a napriek nevýhodam pre náš druh napokon presadil a stal sa preň výhodným. Ak sa to stalo, tak nie preto, že to bolo výhodné pre symbionta Homo sapiens, ale pre symbionov v jeho hlavei, ktorí sa vyvinuli nezávisle od našej evolúcie cez naše mozgy. Výhodné to bolo primárne pre nich, a až následne pre nás.
Rozbor toho AKO a KTORÉ entity je však iba na začiatku. Zatiaľ ten iný živý svet málo chápeme, pretože z tohoto pohľadu to doteraz nikto neštudoval. Ide o úplne iný paradigmatický pohľad, tak nový, že vlastne podobný paradigmatický posun v dejinách asi ani nepoznáme.
Dá sa však hypoteticky predpokladať, že najprv vznikli funkcie — živé bytosti v mozgoch —, ktoré nikto nedelegoval, nemusel sa ich vzdať, ale tieto osídlili niekoho mozog. Napríklad tak, že bol pamätníkom rôznych udalostí a túto pamäť odovzdával svojim potomkom. Mohla takto vzniknúť deľba práce, pretože iní sa chodili pýtať tejto vývojovej vetvy mozgových bytostí. Kým náčelníkov nemajú mnohé lovecko-zberačské kmene (napríklad !Kung Sanovia), tak rôzne vedmy, či šamanov majú skoro pravidelne všetci. Tým sa však vytvoril nový umweltový znak symbiózou umweltov nejakých mozgových bytostí, a to znak výlučnosti šamana. Tento znak má potom šancu byť základom iných takýchto symbióz v usadlých spoločnostiach, má totiž šancu na šírenie cez susedné usadlosti. Možno takto začala špecializácia deľby práce. Ktohovie. Treba skúmať z takéhoto možného uhlu.
- Markoš, A. (2016). Evoluční tápání: Podoby planetárního životopisu. Pavel Mervart 2016.
- Johnson, A. W., & Earle, T. K. (1987). The evolution of human societies: From foraging group to agrarian state. Stanford: Stanford University Press.